Previous Issues Volume 1, Issue 1 - 2017
Histomorphological Developmental Studies of the Left Ovary in the Japanese Quail (Coturnix Coturnix Japonica)
Gamal K Mohamed1 , Aziza A Selim1 , Hanan H Abdelhafeez1 *, Marwa B Mohamed2
1Department of Anatomy and Histology, Faculty of Vet. Medicine, Assuit University, 71526, Egypt. 2Department of Histology, Faculty of Veterinary Medicine, New Valley University, Assuit University, Egypt. Corresponding Author: Hanan H Abdelhafeez, Department of Anatomy and Histology, Faculty of Vet. Medicine, Assuit University, 71526, Egypt Tel: +20 109 144 9737; E-Mail: [email protected]
Received Date: 28 Mar 2017 Accepted Date: 02 Jun 2017 Published Date: 07 Jun 2017
Copyright © 2017 Abdelhafeez HH
Citation: Mohamed GK, Selim AA, Abdelhafeez HH and Mohamed MB. (2017). Histomorphological Developmental Studies of the Left Ovary in the Japanese Quail (Coturnix Coturnix Japonica). Mathews J Cytol Histol. 1(1): 002.
ABSTRACT
72 Japanese quail embryos (Coturnix coturnix japonica) were collected during 3, 5, 7, 9, 11, 13, 15, 17 days of incubation, and one day after hatching and were used for studying the development of the ovary. At 3 day, the indifferent gonads appeared as bilateral cellular thickening located ventromedial to the mesonephros. At this stage, extragonadal germ cells (tissue germ cells) which measured([13, 74), in diameter could be seen migrating in the dorsal mesentery of the hindgut. They had an amoeboid shape. Also circulating germ cells could be demonstrated within the intraembryonic blood vessels near the genital ridges, and had a small diameter about 11.20μm. At 5 day, the left gonads were larger than the right one. These gonads were indifferent, not distinguished into ovaries or testis morphologically or histologically, both consisted of an outer surface epithelium covered a central core of epithelial cell cords.It appeared as a bud-like extension from the germinal epithelium and, was surrounded by the mesenchymal tissue; the prospective stroma of the developing gonad. Few primordial germ cells could be demonstrated between the surface epithelial cells. Macroscopic and Histological sex differentiations were demonstrated at the seventh day of incubation. Left ovary showed a primitive cortex and a primitive medulla. Most germ cells of ovarian cortex were small oogonia beside the few large primordial germ cells. Mitotic figures were frequently demonstrated at this stage. The ovarian medulla was formed of medullary cords, blood vessels and lacunar channel system lined by cuboidal and flat cells and few interstitial cells. At 9 day, the cortex and the medulla were separated by thin layer of connective tissue. The cortex was divided into numbers of the lobe-like structure by surface epithelial invaginations. At 11-17 day the quail ovary was well-organized into an outer cortex and inner medulla. At the day 11, meiotic germ cells were first detected only in the left ovarian cortex.The ovarian cortex was characterized by the presence of two types of germ cells: oocytes located in the central region and oogonia that present in aggregates mainly in the extremities of the ovary, besides to the epithelial cells. At 13 day, interstitial cells were arranged in small cords or groups or single cells in the medulla. At 15 day, the left ovarian cortex was characterized by the presence of cell nests and more interstitial cells with lipid droplet arranged in small groups. The tunica albuginea was formed. The first evidence of primordial follicle formation was demonstrated in the left ovarian cortex of the quail at one day after hatching. The number of germ cells was reached the maximal value at 15th day of incubation declined gradually and reached a minimal value at one day after hatching. The present study showed that 70% of the peak numbers of germ cells was survived in the left ovary of the quail.
KEYWORDS
Ovary; Quail; Embryos; Development; Histochemistry.
INTRODUCTION
The Japanese quail belongs to the order Galliformes, genus Coturnix, and species japonica. [66, 91, 100] The short life cycle, the high fecundity and adaptability to life in cages, the low maintenance cost, and the easy ways to raise and handle it, make the quail an ideal model for research. Moreover, quail production throughout the world is sharply increased for both egg and meat consumption [13, 74]. Regarding the previous features, the Japanese quail is considered an excellent and economic source for research and for animal protein, especially in the developing countries. All these facts place quail in the focus of research and require more information on its reproduction. The ovary is directly involved in the biological economy of the bird and the reproductive ability of sexually mature female birds is dependent on the capacity of the ovary to produce a large number of ova. Although the histological structure of the ovary in different avian species has been studied by several authors [18, 19, 23, 30, 31, 36, 47, 69, 79] in chick embryo, in Indigenous Mallard Duck (Anas platyrhynchos) [6, 101] in Muscovy duck embryos, [47, 76, 80, 88] in quail embryo; [55] of ovaries in ostrich (Struthio camelus) embryo and [77] in ovaries of prey birds . The morphological changes of the germ cells during the different stages of the ovarian development in birds are scanty in the available literature. Moreover, references about the fluctuations in the population of the germ cells during ovarian development in quail are few. The aim of the present study is to give more information on the development of the ovary of the quail using light and scanning electron microscope.
MATERIALS AND METHOD
samples The study using fertile quail (Coturnix coturnix japonica) eggs obtained from the Research Quail Farm connected to the faculty of Agriculture, Assiut University. The fertilized eggs were incubated in a c10 “POULTRY TECHNICAL OFFICE, Alexandria, Egypt” at 37.5°Cwith a relative humidity of 65%. The eggs were rotated automatically every 6 hours after the 3rd day of incubation. We collected 72 quail embryos at day (3, 5, 7, 9, 11, 13, 15, and 17) of incubations. Embryonic stages were determined upon the onset of incubation, in addition to quail’s ovaries at 1st-day post hatching (Figure 1).
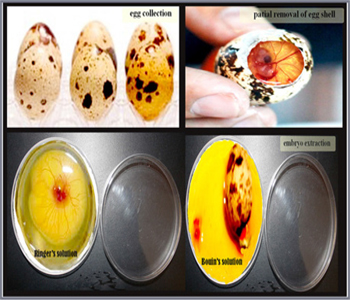
Figure 1: Extraction of embryos.
Isolation of living Embryo Egg shells were opened at the broad end and the shell membranes were carefully removed. Apparent healthy embryos were excised from their shells, placed on a small petri dish, and washed with Ringer’s solution (Figure 2). Small embryos aged 5, 7, 9 and 11 days of incubation were kept at -20°C for 4 hours prior to collecting. Embryos at day 13, 15 and 17 were euthanized by decapitation.

Figure 2: Different stages of quail embryos
Histological preparation Preparation of paraffin sections All embryos were immediately fixed in Bouin’s.Fixed samples were dehydrated in ascending grades of alcohols at 70%, 80%, 90% and 100%. The sampled were cleared using methyl benzoate. Dehydrated samples were then impregnated and embedded in Paraplast (sigma Aldrich). Serial sections of 3-5µm were cut using a Richert Leica RM 2125 Microtome, Germany and mounted on glass slides. Sections were kept in an incubator at 40°C for dryness. The samples were processed according to the following table (table 1). The paraffin sections stained with the several conventional stains including Harris’s Haematoxylin and Eosin for general histological examination [41], Periodic Acid Schiff (PAS) technique for demonstration of neutral mucopolysaccharide [61],Alcian blue 8GX (pH 2.5) [86] counterstained with Harris haematoxylin for demonstration of acidic mucopolysaccharide, Gomori’s reticulin technique for demonstration of reticular fibers [35], Weigert’s resorcin fuchsin for demonstration elastic fibers [102].The stained sections were examined by Leitz Dialux 20 Microscope. Photos were taken using a Canon digital camera (Canon Powershot A95).
| Age/Process | 3days | 5days | 7days | 9days | 11days | 13days | 15days | 17days | 1day post hatching |
|---|---|---|---|---|---|---|---|---|---|
| 1-Fixation | 6h | 8h | 13h | 15h | 17h | 22h | 22h | 5h | 5h |
| 2-Dehydration | |||||||||
| 70% Alcohol I | 2h | 2h | 7h | 3h | 3h | 2h | 2h | 2h | 5h |
| 70% Alcohol II | 1d | 1d | 1d | 2d | 2d | 7h | 12h | 12h | 6h |
| 70% Alcohol III | |||||||||
| 80% Alcohol | 2h | 2h | 2h | 2h | 12h | 12h | 12h | 12h | 12h |
| 90%Alcohol | 2h | 2h | 2h | 2h | 3h | 3h | 3h | 3h | 3h |
| 100% I Alcohol | 1/2h | 1/2h | 1/2h | 1/2h | 3/4h | 1h | 1h | 1h | 1h |
| 100% II Alcohol | 1/2h | 1/2h | 1/2h | 1/2h | 3/4h | 1h | 1h | 1h | 1h |
| 3-clearing with methyl benzoate | |||||||||
| MB I | 2d | 2d | 2d | 2d | 2d | 2d | 2d | 2d | 2d |
| MB II | 2d | 2d | 2d | 2d | 2d | 2d | 2d | 2d | 2d |
| 4embedding in paraffin | |||||||||
| P I | 2h | 2h | 2h | 2h | 2h | 2h | 2h | 2h | 2h |
| P II | 2h | 2h | 2h | 2h | 2h | 2h | 2h | 2h | 2h |
| P III | 4h | 4h | 6h | 18h | 18h | 22h | 22h | 22h | 22h |
Table 1: The processing time of the samples in paraffin embedding techniques. Abbreviation: h, hours; d, days; MB I, methyl bonzoate1, MB II, methyl benzoate II; PI, paraffin I; P II, paraffin II; P III, paraffin III.
Preparation of semithin sections Representative specimens of ovarian tissue were taken from the ovaries of quail embryos at different ages. These samples were immediately fixed in Karnovsky solution [54] at 4°C overnight. Subsequently, the specimens were washed (3 x 15minutes) in 0.1 M sodium phosphate buffer at (pH 7.2). All samples were then post-fixed in 1% osmium tetroxide at 4°C for 2 hr in the dark. Later, they were again washed (3 x 20 minutes) in 0.1 M sodium phosphate buffer (pH 7.2-7.4). Dehydration was performed through graded ethanol to propylene oxide. Samples were dehydrated in ascending graded series of ethanol (50 % (for 30 minutes), 70 % (overnight), 90 % (for 30 minutes), and 100 % I (for 30 minutes) and 100% II (60 minutes). The dehydrated samples were embedded in resin (Epon-Araldite) as the following; Propylene oxide (Merck, Darmstadt, Germany) for 30 minutes, Epon: Propylene oxide about (1:1) (for 30 min) then in Epon (for 3 hours). Epon was prepared (5ml Epon812 (Polysciences, Eppelheim, Germany) + 5ml Araldite + 12ml DDSA) Epon was mixed thoroughly by shaker Incubation at 60 C°. Polymerization of samples was performed by using Epon mix and accelerator (DMP30) (1.5%). The blocks were incubated for 3 days at the following; 60 C° at first day and at 70 C° in the second day and 75C° at third day. Semithin sections (1 µm) were cut using an ultramicrotome Ultracut E (Reichert-Leica, Germany) and stained with toluidine blue (Sodium tetraborate (borax) 1 gram, toluidine blue gram, and Distilled water 100 ml.s) [12]. Scanning electron microscopy Scanning electron microscopy was used to study the epithelial surface of the ovary in the late embryonic stage. The small samples were taken from ovaries at different embryonic ages: 11, 13, 15, 17 days. Samples were washed with normal saline then fixed in Karnovsky fixative composed of (10 ml paraformaldehyde 25%, 10 ml glutaraldehyde 50%, 50 ml Phosphate buffer and 30 ml DW) [54] for 4 hours at 4 C°. Thereafter, they were washed in the Na-phosphate buffer, pH 7.3( 15 min x 4 times )and post-fixed in 1% osmic acid in 0.1 M Na-phosphate buffer for further 2 h at room temperature. Thereafter, they were washed by 0.1 M Na-phosphate buffer15 min x 4 times. The samples were then dehydrated by alcohol 50%, 70%, 90% for 30 min in each concentration and 100% for 2 days with changes many times followed by isoamyl acetate for 2 days. Then subjected to critical point drying method with a (Critical Point Drying Procedure Polaron E3000 CPD Apparatus, Germany). Finally, they were coated with gold using (JEOL -1100 E-ion sputtering Device, Japan) and observed with JEOL scanning electron microscope (JSM– 5400 LV, Japan) at KV 10 at the Electron Microscopy Unit of Assiut University.
Morphometrical measurements The morphometrical studies were applied on paraffin and a semithin section of representative areas of ovaries at various stages of development using image tool 3 computerized program for measuring and counting the number of germ cells per unit area. The quantitative data was presented as mean ± SE.
RESULTS
Gross morphology of ovarian development in quail embryos Macroscopic observation showed that the two ovaries became differentiated at 7 days of incubation, they were located ventromedial to the corresponding mesonephros, the left ovary was larger and somewhat dorsal to the right one. Moreover, the left ovary was located somewhat cranial to the right one. They were converged caudally to become in contact with each other at their caudal end (Figure 3A). At 9days of incubation macroscopic observation showed the clear asymmetry between left and right ovaries (Figure 3B). At 13 days of incubation, the left ovary became very large in comparison with the right one, it was a bean in the shape its medial border was convex while the lateral border was concave. Its cranial extremity which directed laterally and cranially was larger than the caudal end which directed laterally and caudally, the apex of the convexity of the medial border of the left ovary came in contact with the caudal end of the right ovary.The right one was represented by the thin slender structure, its longitudinal axis was directed caudally and medially.It was located on the ventral aspect of the junction of the proximal fourths of the right kidney. Left one occupying the ventral aspect of the proximal three-quarters of the medial part of the kidney (Fig. 3C). At 17 days of incubation, the left ovary was very large in comparison to the right one , the left one was a bean-shaped its medial border was convex while the lateral border was concave (Figure. 3D) . At 1day post-hatching, the left ovary was situated in the peritoneal cavity related to the anterior end of the kidney and the caudal end of the lung. It was whitish or creamy in color elongated (three times as long as wide) with a deep groove separated the cranial and caudal parts of the ovary or it could be seen shorter, broader and more elongated at the cranial end, while the right ovary was barely visible after hatching (Figure 3E and F)
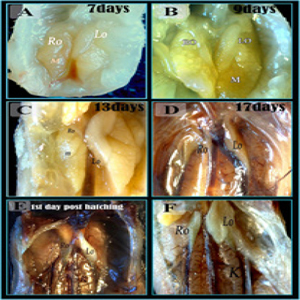
Figure 3: Gross morphology of ovarian development in quail embryos.
Photographs of right and left ovaries in the abdominal cavity in 7 days (A), 9 days (B), 13 days (C), 17 days (D) of quail embryos and 1-day post-hatching (E, F). Note: Left ovary (LO), Right ovary (RO), mesonephros (M), kidney (K).A: the two ovaries became pormenant. Note, Left ovry (LO), right ovary (RO). B: clear asymmetry between left and right ovaries Note, Left ovry (LO), right ovary (RO)., mesonephros (M). C: showed the asymmetry between left functional ovary (LO) and the right regressing ovary (RO). D: the left ovary (LO) was very large in comparison to the right one (RO), the left one was a beanshaped its medial border was convex while the lateral border was concave. E, F: The left ovary was situated in the peritoneal cavity related to the anterior end of the kidney (K).
MICROSCOPIC OBSERVATIONS
Quail embryo at 3 days of incubation The indifferent gonads at 3 days of incubation could be identified as a bilateral cellular thickening located ventromedial to the mesonephros (Figure 4A, B). The left genital ridge was formed of somatic cells, few numbers of primordial germ cells and covered by the coelomic epithelium (Figure 4C). The gonadal coelomic epithelium was formed of crowded oval or elongated epithelial cells their cytoplasm was deeply stained basophilic and contained darkly stained nuclei, While the primordial germ cells were distinguishable by their large diameter and it mean diameter was measured about (13.79μm), spherical or oval shape and their cytoplasm was faintly stained and vacuolated contained prominent nuclei, measured about (8.55μm) (Figure 4C and H). Germ cell could be aligned with the surface epithelium (Figure 4E and G). Extragonadal germ cells migrated in the dorsal mesentery (Figure 4D). Extragonadal germ cells could be seen migrating in the dorsal mesentery of the hindgut (Tissue germ cells); they had amoeboid in shape (Figure 4G). Tissue germ cells were in early stages of active mitotic division Metaphase (Figure 4 G and I) and Anaphase was demonstrated (Figure 4F).Semithin section stained with toluidine blue showed that the nucleus of primordial germ cells was in the active mitotic division (Figure 4I). Also circulating germ cells could be demonstrated within the intraembryonic blood vessels near the genital ridges; these circulating germ cells had a smaller diameter than those of the gonadal germ cells their mean diameter was measured about (11.20μm) (Figure 4J, K, L). The primitive gonads showed a negative reaction to both PAS and Alcian blue stains. Paraffin section (A, B, C, D, E, F) stained by HE and semi-thin sections stained by toluidine blue (G, H, I, J, K,L) of 3 days old quail embryo .A: the arrows refer to genital ridges, Mesonephros (M), Neural tube (Nt), Aorta (AO). Note located ventromedial to the mesonephros B Showing the the genital ridges (arrowheads), Aorta (AO), Mesonephros (M), Notcord (N), Nephric duct (Nd), Mesentery (Ms). C: Higher magnification of fig B showing that the left genital ridge was formed of somatic cells (arrowhead) and primordial germs cells (arrows). D: extragonadal germ cells migrating in the dorsal mesentery. Tissue germ cell showing metaphase stage of mitotic division (T), PGC at the coelomic angle (A), PGC in the genital ridge (G), Mesentery cells (Mc), Genital ridge (GR). E: showed somatic cells (arrowhead) and primordial germ cells (arrow). F: showed primordial germ cells in mitotic division (arrow).G: migrating germ cells (arrows), germ cell had ameboid shape (arrowhead). H:Mesothelial lining of genital ridge(arrow heads) and Mesenchymal cells (Mc). Note : invading primordial germ cells(double arrow heads). H: mesenchymal cells(MC) and primordial germ cells (arrowheads) in mitotic division. J: masses of migrating germ cells (arrows) inside the blood vessel and celomic cavity. Note arrowhead refer to the genital ridge. K: Higher magnification of the intravascular primordial germ cells(PGCs),endothelial linning of the vessel(arrowhead).L: The invading primordial germ cells( (PGC) were invading the endothelium of blood vessels (white arrowhead,PGC). Note: RBCS and the ameobid features of primordial germ cells(whitearrowheads).
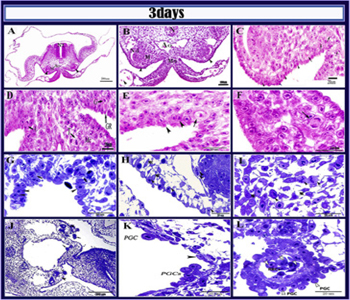
Figure 4: Development of the genital ridge.
Quail embryo at 5 days of incubation The genital ridges at 5 days of incubation were slightly increased in size and bulged into the coelomic cavity as distinct organs located on the ventromedial aspect of the mesonephros. The gonads at this stage were small in size located on the either side of the mesentery ventral to the aorta. In cross section, they appeared spherical-shaped and there was an asymmetry between left and right gonads, where the lef was slightly larger than the right one (Figure 5A, B). At this embryonic stage, these gonads were indifferent, not distinguished into ovaries or testis morphologically. Both consisted of an outer surface epithelium covered a central core of epithelial cell cords (Somatic cell cords). Surface epithelium was formed of cuboidal or pyramidal cells. Gonadal germ cells located mainly at the periphery while few located deeper in the cortex. They were large in size, spherical in shape, with large vesicular nuclei, and faintly stained vacuolated cytoplasm (Figure 5C, D, E). The germ cells were undergoing meiotic division and were observed in different stages; prophase (Figure 5F), metaphase (Figure 5H), anaphase (Figure 5G). The epithelial cell cords appeared as a bud like an extension from the germinal epithelium, surrounded by mesenchymal tissue to form the stroma of the developing gonad (Figure 5C). Gonadal stromal fibers and outer thin coat of the surface epithelium gave a positive reaction with PAS (Figure 5J) and Alcian blue (Figure 5I). Moreover, fine reticular fibers supported the gonads (Figure 5K).
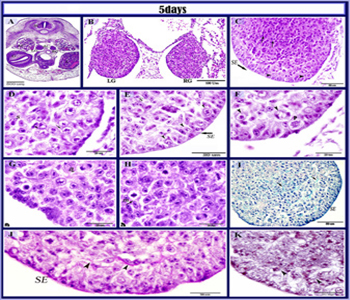
Figure 5: Histological structure of the indifferent gonads in 5 days old quail embryos.
paraffin section in 5 days old quail embryo stained with HE (A, B, C, D, E, F, G, H), Alcian blue (I), PAS (J), Gomori’s reticulin stain (K). A, B: showed the position of the gonads (LG) Left gonad, (RG) Right gonad, (M) mesonephros, (L) liver, (G) gizzard, (sc) spinal cord. A, B: Note. the asymmetry between the indifferent gonads. The gonads were slightly increased in size and bulged into the coelomic cavity and located on the either side of the mesentery ventral to the aorta (Ao). C: Showed outer surface epithelium (SE,arrow), epithelial cell cords (arrows) and germ cells (arrowhead). D: showing that the primordial germ cells (arrows) were localized at the periphery, (S) somatic cells. E: The cuboidal or pyramidal surface epithelial cells (SE), some germ cells interposed among surface cells (arrowhead) ;(s) somatic cells (S). F, G, H: gonad showing germ cells at different stages of mitotic division anaphase (a,arrowhead), metaphase (m,arrowhead). prophase (p), germ cells (arrowheads). I: positive AB stromal fibers(arrows) and outer thin coat of the surface epithelium (SE). J: positive PAS outer thin coat of the surface epithelium (SE) and positive stromal fibers (arrowhead) (Periodic Acid Schiff reagent). K: fine reticular fiber within the stroma (arrowhead).
Quail embryo at 7 days of incubation Histological cross section showed the right one was more oblique in its position than the difference between the left and right ovaries in size, shape and structure difference in size between the two ovaries was more obvious than that observed in the previous embryonic stage (Figure 6A and B). At this stage of development, histological sex differentiation could be demonstrated. The general architecture of the left ovary could be distinguished into the primitive cortex and a primitive medulla (Figure 6C). The ovarian cortex was covered by surface epithelium and contained germ cells and somatic cells (Figure 6D). Germ cells at this stage were primordial germ cells in the active mitotic division besides to oogonia; no signs of meiosis could be demonstrated in the germ cells (Figure 6D and E). The resulting daughter cells (oogonia) were smaller than the primordial germ cells their diameter was measured about (9.2μm), rounded or oval in shape their cytoplasm was clear contained large vesicular nucleus contained 1-3 distinct nucleoli. These germ cells were arranged singly in the cortex, normal oogonia comprised the most germ cells population while apoptotic germ cells were rarely demonstrated (Figure 6E and F). The outer surface epithelium was formed of columnar or cuboidal cells, that arranged in single layer mainly, but it could be multilayered in other regions (Figure 6F and I). Some of the ovarian surface epithelial cells could be demonstrated in the mitotic division. Some primordial germ cells were demonstrated among the surface epithelial cells in the active mitotic division to form oogonia (Figure 6F). The Ovarian medulla of the left was formed of irregular disorganized cords of epithelial cells (the medullary cords), blood vessels and distended medullary cords lined with flattened or cuboidal cells (Lacunar channel system). Few interstitial cells could be demonstrated (Fig 6G and H). These cells were oval, rounded or polyhedral cells, their nuclei were vesicular and eccentric, their cytoplasm was pale acidophilic and vacuolated. Also, there were medullary germ cells characterized by a condensed chromatin similar to those of meiosis demonstrated among the surface epithelial cells in the active mitotic division to form oogonia (Figure 6H). The outer surface of the left ovary showed PAS (Figure 6I and J) and alcian blue (Figure 6K and L) positive coat. PAS (Figure 6J) and alcian blue (Figure 6L) Positive fibers were observed in ovarian stroma.
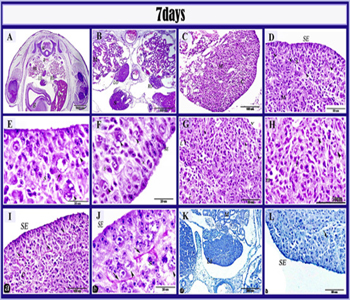
Figure 6: Histological structure of the left ovary in 7 days old quail embryos.
Paraffin section in 7 days old quail embryo stained by HE (A,B, C, D, E, F, G,H), PAS (I, J), alcian blue (K, L). A, B: showed the asymmetry between left ovary (LO) and right ovary (RO), liver (L), mesonephros (m), gizzard (g), aorta (a), and spinal cord (sc). C: the left ovary formed of the cortex (c) and medulla (M). D: the cortex contained germ cells (arrowheads) and somatic cells (s), medulla (M), surface epithelium (SE). E: oogonia (arrowhead), primordial germ cells (arrow), somatic cells (S). F: germ cell (A) interposed among surface epithelial cells in anaphase stage (A) of mitotic division, other germ cells located in the cortex (arrowheads), somatic cells (s) and surface epithelium (SE) Note: bulging structure on the surface epithelium. G: ovarian medulla was contained mesenchymal cells (m), lacuna (L), germ cells (arrowhead), interstitial cells (I) and blood vessels (v).H: Higher magnification of medulla showing the interstitial cells (I), germ cells (arrowheads). I, J: PAS positive stromal (arrows) fibers and thin outer surface epithelial coat (SE), germ cells (arrowhead), somatic cells (S), surface epithelium (SE). K, L: Ab positive stromal fiber(arrows) and Ab positive outer thin coat of the surface epithelium (SE).Note:L ,left ovary and M,mesonephros.
Quail embryo at 9 days of incubation Histological cross section showed that the left ovary was larger about four times than the right one, due to its enlargement, it was migrated medially to become located ventral to the aorta and medial to the mesonephros. On the other hand, the right remains smaller and on its early position ventromedial to the mesonephros (Figure 7A, B). The cortex and the medulla of left ovary were clearer and separated by thin layer of connective tissue (Figure 7F). The cortex became thicker than that of the previous age as the cortical cells were proliferated and increased in number arranged in aggregates or irregular cords separated by a connective tissue strands (Figure 7C, D). Germ cells that showed active mitotic figures were demonstrated at this stage and increased in amount, oogonia in prophase, metaphase more frequent than other stages (Figure 7E, H). The inner part of the cortex was relatively loose than the outer part also the cortical tissue at the margin of the medulla showed an extension downward into the medullary tissue. The surface of the cortex showed different degrees of invaginations that divided the cortex into numbers of lobes-like structure (Figure 7C). The ovarian cortex was covered by outer surface epithelium with some germ cells were interposed among surface epithelial cells (Figure 7D and E). There were clusters of germ cells formed of (3-4) cell found on the deep surface of the epithelium that forms lobulation, these buds form the cortical cords (Figure 7C, D). The medullary tissue showed a massive increase in size. The medullary cords were well-organized and became longer and thicker. The medullary cords located between the wide lacunae.Germ cells could be distinguished in the medulla. Some germ cells undergo degeneration (Figure 7F and G). The reaction of the stromal fibers to PAS (Figure 7I) and Alcian blue (Figure 7J) became stronger than the previous stages. Few elastic fibers could be demonstrated in the ovarian cortex by Weigert’s technique (Figure 7K).
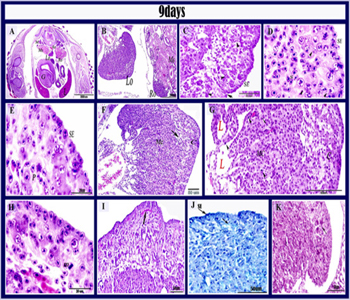
Figure 7: Histological structure of the left ovary in 9 days old quail embryos.
Paraffin section in 9 days old quail embryo stained by H&E (A, B, C, D, E, F, G, H), PAS (I), Alcian blue (J), Weigert’s stain (K). A: showed the position of left and right ovaries in the body cavity.the right ovary was smaller and keep its position in ventromedial to the mesonephros (Ms), liver (L), gizzard (G), left ovary (LO), right ovary (RO), Aorta (Ao), Metanephros (M). B: showed the asymmetry between left and right ovaries, mesonephros (Ms) left ovary (LO); right ovary (RO). C, D: The cortex contained aggregates of oogonia (arrowheads), surface invagination (arrow), surface epithelium (SE) formed of cuboidal or columnar cells, somatic cells (S). E: cuboidal cells of the surface epithelium (SE), germ cells in mitotic division (Prophase) (P). F: showed the medullary cords (Mc), cortex (c) and primitive connective tissue layer (arrow). G: the medullary tissue contained lacuna(L), medullary cords (Mc), degenerated germ cells (arrowhead), normal germ cells (arrow) and cortex (C). H: showed germ cells in mitotic division (metaphase) (m). I: showed PAS-positive stromal fibers (arrow). J: showed that the outer thin coat of the surface epithelium positive to Ab (SE). K: showed few elastic fibers in the ovarian cortex (arrow).
Quail embryo at 11 days of incubation The asymmetry between the left functional ovary and the right regressing ovary was still clear (Figure 8A, B). The cortex had somatic cells, several stages of the germ cell cycle could be observed such as oogonia, oocyte in different stages of meiotic divisions (Figure 8C, E, F, and G). Zygotene stage was more frequently demonstrated at this stage. Oocyte at zygotene was characterized by thick, shortened chromosomes (bouquet formation) and bead-like structure along the whole length of the chromosomes was occasionally demonstrated (Figure 8G, H and I). The pre-meiotic resting oogonia existed in aggregates mainly in the extremities of the ovary (Figure 8E and F). The medulla of the left ovary was increased in size and it was formed of two layers, outer compact layer which lied next to the cortex and composed of condensed solid medullary cords and cells, the inner reticular layer was formed of medullary cords separated mesenchymal by strands of connective tissue, blood vessels, and lacuna. Medulla became more vascularized, had a well-developed lacuna, and Degenerated germ cells were found attached to the epithelial lining of the lacunar channels or released in the lacuna singly or in masses of 3-4 cells. (Figure 8D, J, and K). The interstitial cells were increased at this stage could be seen in groups or nest surrounded by a connective tissue (Figure 8D and G).The surface epithelium was multilayered (SE) which gave rise subepithelial cords (Figure 8I). The connective tissue stroma of the cortex became more obvious. With a Periodic Acid Schiff (Figure 8L) andAlcian blue stains (Figure 8N) the stroma showed positive reaction stronger than the previous stages and a PAS positive coat surrounded the surface epithelium. With Gomori's reticulin technique stroma showed positive reticular fibers (Figure 8M) and elastic fibers with Weigert’s technique.
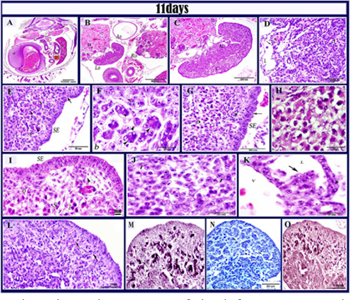
Figure 8: histological structure of the left ovary in 11 days old quail em- bryos.
Paraffin section in 11 days old quail embryo stained by H&E (A,B, C, D, E, F, G, H, I, J, K), PAS (L), Gomoris reticulin stain (M), alcian blue (N), (O) Weigert’s elastic. A: showed the position of two ovaries in the abdominal cavity left ovary (LO), right ovary (RO), mesonephros (m); liver (L) and gizzard (g) B: higher magnification of fig (A) showed the asymmetry between left and right ovaries Left ovary (Lo), Right ovary (Ro), Mesonephros(M), Aorta (Ao). C: showed cortex (c); medulla (m). D: the medullary cords (mc), blood vessels (v), Lacunar channels (L), degenerated germ cells (arrow) and groups of interstitial cells (I). E: the extremity of the cortex contained oogonia (arrowhead) and somatic cells (s) and surface epithelium (SE). (F) higher magnification fig (E) note; oogonia (arrowhead), somatic cells (s). G,H: the cortex at the central part contained oocyte in meiotic prophase somatic cells (s) oocytes (O) in zygotene stage at first meiotic prophase (z). Note: subepitheila cords (arrowhead). I: the surface epithelium was multilayered (SE) which gave rise subepithelial cords (arrowhead). Note oogonia (o), oocytes in zygotene (z), somatic cells (s). J: the interstitial cells (I) located in the medulla .K: Degenerated germ cells (arrow) in the medullary lacuna (L) attached to epithelial lining of the lacuna and blood vessels (V). L: positive PAS stromal fibers (arrows). M: fine reticular fibers in the ovarian stroma (arrow). N: Alcian blue positive stromal fibers(arrows)(O): positive Weigert’s stromal fibers (arrows).
Quail embryo at 13 days of incubation In cross section, the left ovary appeared to be located more dorsally to become in contact with the aorta, this because its extension at the expense of the area that became free due to degenerated mesonephros (Figure 9A). The left ovary was characterized by marked the thickness of the ovarian cortex. The cortical cords were increased in size and number and extended into the medulla, containing more germ cells and somatic cells than in the previous ages (Figure 9B and C). The germinal epithelium had several layers of columnar cells which were supported by a primitive tunica albuginea. The surface epithelial cells sometimes appeared to extend downward in the cortex to form subepithelial cords (Figure 9E and F). Ovarian cortex was predominated by ovarian germ cells which could be classified into two groups the first group, oocytes in the prophase stages of the first meiotic division. (leptotene, zygotene) were demonstrated at this embryonic stage mainly in the central region of the ovarian cortex (Figure 9C and D). The second group of cortical germ cells was oogonia at resting pre-meiotic phase mainly at the two extremities of the cortex and mitotic division could be seen (Figure 9H and L). The ovarian surface invaginated into the cortex (Figure 9 F). The ovarian medulla also increased in size as a result of the formation of additional medullary cords and distension of the others. It was more vascular ized than the previous age (Figure 9 G, H, M). The medulla of left ovaries presented the same components as in the previous age but presented a laxer tissue with more germ cells forming clusters, more developed lacunar channels and more interstitial cells were arranged in small cords or groups and some single interstitial cell) Figure 9M and I). Degenerated germ cells could be observed in the lacunar channels (Figure 9N). Stromal fibers gave a positive reaction with Periodic Acid Schiff (Figure 9J and K) and with Alcian blue (Figure 9O) Ovarian stroma had fine reticular fibers that appeared black with Gomori’s reticulin technique (Figure 9P).
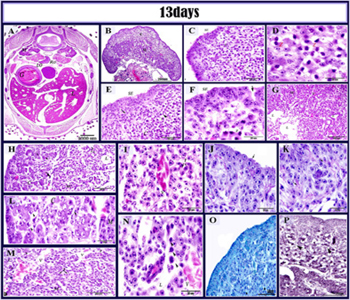
Figure 9: histological structure of the left ovary in 13 days old quail embryos.
paraffin section in 13-day old quail embryo stained by HE (A, B, C, D, E, F, G, H, I, L, M, N), PAS (J, K), Alcian blue (O), Gomori’s reticulin stain (P). A: showed the position of the left and right ovary (M) metanephros ;(LO) left ovary ;(RO) right ovary ;(G) gizzard and (L) liver. B: cortex (c) and medulla (m) C: showing cortex contain; oocytes in zygotene stage (z) and somatic cell (s) Note that surface epithelium was formed of multilayered (SE), subepithelial cords (arrow).D: oocytes in zygotene (z). E: surface epithelium was formed of simple columnar epithelium or multilayered (SE), subepithelial cords (arrow), cortex (c), oocytes (arrowhead) and somatic cells (s). F: A higher magnification showing multilayered surface epithelial cells (SE), Somatic cells(s) and oocytes in zygotene stage (z), surface epithelium invagination (arrow). G: ovarian medullary cords (MC), blood vessels (V) and lacuna (L). H: cortex (c), germ cells (arrowhead), primitive tunica albuginea (arrow), medullary cords (MC), lacuna (L) and interstitial cells (I). I, M: Higher magnification showing that medulla was formed of lacuna (l), interstitial cells (I), blood vessels(V).L: cortex contain oogonia at pre-meiotic resting phase(arrowhead), surface epithelium (SE), Medulla (M), Cortex (C). J: PAS positive outer coat of the surface epithelium (arrow). K: PAS positive stromal fibers (arrows). N: the medulla showing degenerated germ (arrowhead) within the lacunar channels (L), Blood vessel (V). O: the alcian blue positive outer coat of the surface epithelium (SE). P: distribution of the reticular fibers (arrowhead).
Paraffin section in 11 days old quail embryo stained by H&E (A,B, C, D, E, F, G, H, I, J, K), PAS (L), Gomoris reticulin stain (M), alcian blue (N), (O) Wigert’s elastic. A: showed the position of two ovaries in the abdominal cavity left ovary (LO), right ovary (RO), mesonephros (m); liver (L) and gizzard (g) B: higher magnification of fig (A) showed the asymmetry between left and right ovaries Left ovary (Lo), Right ovary (Ro), Mesonephros(M), Aorta (Ao). C: showed cortex (c); medulla (m). D: the medullary cords (mc), blood vessels (v), Lacunar channels (L), degenerated germ cells (arrow) and groups of interstitial cells (I). E: the extremity of the cortex contained oogonia (arrowhead) and somatic cells (s) and surface epithelium (SE). (F) higher magnification fig (E) note; oogonia (arrowhead), somatic cells (s). G,H: the cortex at the central part contained oocyte in meiotic prophase somatic cells (s) oocytes (O) in zygotene stage at first meiotic prophase (z). Note: subepitheila cords (arrowhead). I: the surface epithelium was multilayered (SE) which gave rise subepithelial cords (arrowhead). Note oogonia (o), oocytes in zygotene (z), somatic cells (s). J: the interstitial cells (I) located in the medulla .K: Degenerated germ cells (arrow) in the medullary lacuna (L) attached to epithelial lining of the lacuna and blood vessels (V). L: positive PAS stromal fibers (arrows). M: fine reticular fibers in the ovarian stroma (arrow). N: Alcian blue positive stromal fibers(arrows)(O): positive Wigert’s stromal fibers (arrows).
Quail embryo at 15 days of incubation Left and right ovaries were asymmetric (Figure 10A). The general architecture of the left ovary became more prominent and the cortex and medulla were well-defined (Figure 10B). Cortex was increased in thickness and characterized by the presence of cell nests. Each cell nest contained large germ cells and small rounded somatic cells present mainly in the central part of the ovary. The cell nests were separated from each other by spindle-shaped stromal cells (Figure 10C, D). Most germ cells were oocytes in meiotic prophase, zygotene (Figure 10 I, J and K), leptotene (Figure 10G).; pachytene (Figure 10F). The outer surface epithelium was separated from the e cortex by a layer of connective tissue; tunica albuginea which was rich in germ cells (Figure 10D). Ovarian epithelial cells showed multiple sites of invagination (Figure 10E). Ovarian medulla was increased in thickness than the previous embryonic stages and characterized by enlarged lacunar channel system that occupied the basal portion of the ovary. Dead medullary germ cells were present in groups or masses of 3-4 cells released in the lacuna (Figure 10M, N). There were more interstitial cells than those of the previous stages arranged in small groups with more lipid droplets (Figure 10 O, P). The ovarian stromal fibers were reacted positively to both PAS and Alcian blue stain but the reaction was .With with Wigert’s technique stroma showed positive and elastic fibers (Figure 10L).The organization of the reticular fibers dividing the cortex into lobules while they were interspersed between the medullary cells (Figure 10H).
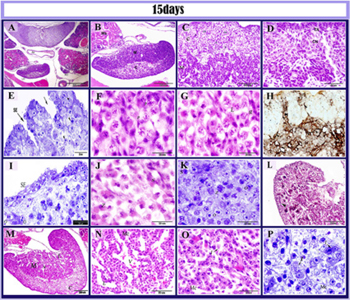
Figure 10: histological structure of the left ovary in 15 days old quail em- bryos.
paraffin (A,B, C,D, F, G, J, M, N, O) stained by HE and semi-thin (E, I, K, P) section in 15 day old quail embryo and semithin sections stained by toluidine blue (E, I K, P), Gomori’s reticulin stain (H), Wigert’s Elstica. A: showed the asymmetry between the ovaries left ovary (LO), right ovary (RO), mesonephros (Ms), aorta (Ao). B: the left ovary consisted of the cortex(C) and medulla (m). C: well-defined cortex and medulla. D: the tunica albuginea (TA), germ cell nest (CN). E: invagination surface epithelial (arrows), note surface epithelium (SE). F: oocytes in a pachytene stage (p). G: oocytes in leptotene stage (L), stroma cells (st). H: showed the reticular fibers (arrowheads) demarcating a lobular-like areas in the cortex while reticular fibers organized in a network (N) in the medulla. I: surface epithelium (SE), oogonia in pre-meiotic resting phase (O), zygotene stage (Z). J: oocytes in zygotene stage (Z), stromal cells (S). K: oocytes in zygotene stage (Z), oogonia in a pre-meiotic resting phase (O) and somatic cells (s). L: positive Wigert’s stromal fibers (arrows). M: more vascularized medulla (M), blood vessels (V), lacunar channels (L). N: Higher magnification of fig (M) showing medullary cords (MC) and blood vessels(v) and lacuna (L). O: Interstitial cells (I), medullary cords (Mc). P: interstitial cells (I,arrows), medullary cord (Mc)
Quail embryo at 17 days of incubation The developmental changes were similar to those of the previous embryonic developmental stages where the ovarian cortex was more developed and increased in thickness contained two types of cells, the somatic cells, and meiotic germ cells. The later organized in groups; the cell nests which were separated by connective tissue layer (Figure 11C). Different stages of meiotic prophase leptotene (Figure 11B and C), zygotene (Figure 11C), pachytene (Figure 11D) and diplotene stages could be demonstrated. Pachytene and leptotene were more frequently demonstrated than other prophase stages (Figure 11A-F). The outer surface epithelium of the ovary was reduced to single or two layers of cuboidal cells (Figure 11E). Surface epithelial cells were appeared to extend into the cell nests and contact the germ cells forming invaginations into the deep cortex and sub-epithelial cords (Figure 11F). Medulla was thin in thickness characterized by the presence of large blood vessels, well-developed lacuna (Figure 11A, G, and H) and more interstitial cells (Figure 11I). paraffin section stained with HE (A,B, C, D, F, G,H) and semithin sections stained with toluidine blue (E,I) in 17-day old quail embryo. A: cortex (c), medulla (m), germ cell nest (arrows)blood vessels (v). B: surface epithelium (SE), oocytes in leptotene (L) , stroma cells (s). C: the cortex contained cell nest (CN), oocytes in leptotene (L), oocytes in zygotene (Z), Somatic cells (S). D: oocytes in pachytene stage (p) and stroma cells (st). E: oogonia (arrows) in a pre-meiotic resting phase. Note. the surface epithelium (SE). G: invagination of the surface epithelium (arrow). H, I: medullary cells (Mc), blood vessels (v). J: medullary interstitial cells (I).
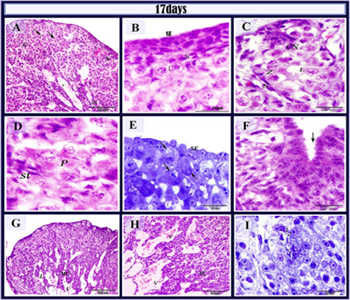
Figure 11: histological structure of the left ovary in 17 days old quail embryos.
Quail ovary, at the 1st-day post-hatching The general organization of the left ovary was well-established into; an outer cortex and inner medulla. The cortex and medulla were separated by a thin connective tissue layer and the two regions could be easily differentiated (Figure 12A and B). The ovarian surface epithelium was formed of one or two layers of flattened cells supported by a connective tissue layer; the tunica albuginea (Figure 12C and E). Occasionally ovarian surface epithelial cells appeared to extend into the cell nests and contact the germ cells forming cortical invagination. These cortical invaginations were increased in both depth and number as compared with those observed at the previous developmental stages (Figure 12C, D). The ovarian cortex contained large clusters of germ cells. Most of these germ cells were oocytes in various stages of meiotic prophase (Figure 12E and F).The first evidence of primordial follicle formation was demonstrated in the left ovarian cortex of the quail at one day after hatching (Figure 12C), While oogonia were markedly reduced in number. Oocytes gathered small follicular cells around them and formed the first primordial follicles (Figure 12G, H and I). The oocyte of the primordial follicle was demonstrated at the resting diplotene stage of the first meiotic prophase and it was large in size with eccentric nucleus contained single nucleolus and diffuses chromatin (Figure 12 I). The medulla of the left ovary was very fragmented with numerous lacuna and more interstitial cells, each cell of these cords was polyhedral or elongated in shape, the nucleus was eccentric, vesicular and contained distinct nucleoli and their cytoplasm was faintly stained and showed vacuoles of differ ent sizes. The lacunar channel system was more developed than those observed in the previous developmental stages (Figure 12A, B, G).
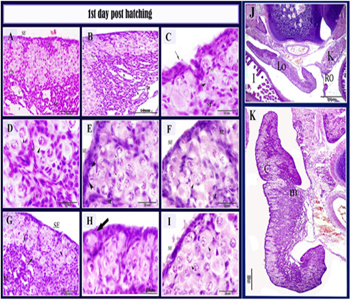
Figure 12: histological structure of the left ovary in 1 days post-hatching quail bird.
Paraffin section stained with HE in 1-day post-hatching quail bird. A, B: the cortex (C), Medulla (M), cell nest (CN), blood vessels (V), Lacuna (L). C: invagination of surface epithelium (arrow), the primordial follicle (PF), stromal cells (St). D: oocytes in groups (arrowhead) surrounded by stromal cells (st), somatic cells (s). D: the surface epithelial cells in closed contact with the germ cell (arrowhead), oocytes in pachytene stage (arrows), stromal cells (St), flattened surface epithelium (SE). F: oocytes in leptotene stage (arrowhead), stromal cells (St), surface epithelium (SE). G, H: primordial follicle (arrow), cortex (c), medulla(m), germ cell (arrowhead). I: oocytes in diplotene stage (arrowhead), surface epithelium (SE), stroma cells (st).(J) : showed the position of the left (LO) and right ovary;(RO); (I) intestine; K (kidney) and the asymmetry between the ovaries left ovary and right ovary.K: : the left ovary consisted of the well defined cortex (C) and medulla (m).
Scanning electron microscopic observation of the quail ovary at the late embryonic stages (11-17) day of incubation Revealed that the normally developing left ovary was spindleshaped and its surface appeared rough at low magnification. The epithelial cells were usually rounded or oval cells but sometimes were fusiform in shape. The surface of each cell was bulging carrying much protrusion such as microvilli or globular projections were present on the cell surface. In 11 days of quail embryos, most of the ovarian surface epithelial cells were fusiform in shape. A globular-like structure was recognized on the ovarian surface (Figure 13A-C). In 13 days of quail embryos, the ovarian surface was irregular and most surface epithelial cells were rounded or oval in shape. globular like structure was observed between surface cells. Microvilli were identified on the surface of the ovarian epithelia (Figure 13D-F). F: showed the ovarian surface epithelial cells. short cilia (Figure 13F). In 15 days of quail embryos, the left ovary had an irregular surface (Figure 13G). A ridge-like protrusion existed on the surface epithelial cells (Figure 13H). A marked invagination was observed on the surface of the ovarian epithelium (Figure 13 I). In the 17 days of quail embryos, the epithelial cells had rounded surface. A ridge-like protrusion attached to the ovarian surface epithelial cells (Figure 13J, K, L).
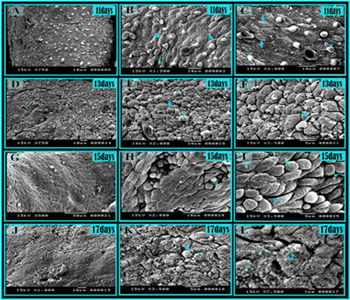
Figure 13: Scanning electron micrograph of the ovarian surface in quail embryos.
Scanning electron micrograph of the left ovary of 11 days (A,B,C), 13 days (D,e, F), 15 days (G,H, ,I), 17 days (J,K,L) .quail embryos. A: general view of showing the irregular surface of the ovary. B, C: Most of the ovarian surface epithelial cells were fusiform in shape (Arrows), Note the globular like structure (arrowhead). D, E:D general view andE higher magnification of the ovarian surface was irregular and most surface epithelial cells were rounded or oval in shape (arrow). Note. globular like structure on the surface cells (arrowhead) .F: showed the ovarian surface epithelial cells. Note single long t cilia (arrow) G: the left ovary had an irregular surface. H: invagination of the surface epithelium (arrowhead) I: higher magnification showed ridge-like protrusion on the surface epithelial cells (arrowheads). J: showed the ovarian surface epithelial cells. K: rounded surface epithelial cells (arrow) ridge-like protrusion on the ovarian surface epithelial cells (arrowheads). L: Higher magnification showed the ridge-like protrusion on the ovarian surface epithelial cells (arrowheads) and microvilli (doublearrows).
Quantification of the germ cells in quail during different stages of embryonic development Quantitative analysis revealed that the germ cell population per unit area was few at an early stage of embryonic development. The number of germ cells was gradually increased with the advancement of embryonic development and reached the maximal value at 15th day of incubation. With the advance ment of development, the germ cells declined gradually and reached a minimal value at one day after hatching (Figure 14 and table 2). The present study showed that in quail only 70% of the peak numbers of germ cells was survived in the left ovary of the quail.
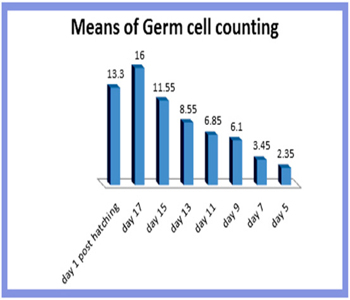
Figure 14: Mean number of germ cell during different stages of ovarian development per unit area (3850,25uµ2) and expressed as means±SE(standerd error).
| Age | Mean number of germ cells |
| 5 days | 2.35± 0.59 |
| 7days | 3.7 ± 1.64 |
| 9 days | 6.3 ± 2.19 |
| 11 days | 8.65 ± 2.8 |
| 13 days | 11.55 ± 3.09 |
| 15 days | 14 ± 2.76 |
| 17 days | 11.8 ± 4.3 |
| 1 day post | 9.5 ± 3.2 |
Table 2: Mean numbers of germ cell during different stages of ovarian development per unit area. (3850,25uμ2) are expressed as means± SE (Standerd Error).
DISCUSSION
The current observations on the histomorphological studies of development of the ovary in quail revealed that the indifferent gonad of the quail embryo at third day of incubation could be identified as bilateral cellular thickening located ventromedial to the mesonephros. Each gonad at this early stage of embryonic development was represented by a thickened coelomic epithelium contained few number of primordial germ cells. The gonadal coelomic epithelium was formed of crowded cuboidal or columnar or elongated epithelial cells, showed no distinct basement membrane. The germinal epithelial cells were numerous and smaller in size compared with those of the primordial germ cells. Their cytoplasm was basophilic and contained darkly stained basophilic nuclei, arranged irregularly on the surface of ceolomic gonadal epithelium. Similar findings were demonstrated by several authors [5, 7, 23, 24, 38, 49, 52, 59, 94]. The formation of the gonads in the female Japanese could be considered as beginning with the arrival of primordial germ cells (PGCs) in the peritoneal epithelium midway between the cranial aspect of the mesonephric ridge and the developing dorsal mesentery. This happened at about 48 to 60 hours incubation and the germinal epithelium was formed of two to three layers and an underlying tissue of loosely arranged mesenchymal cells [52]. Chang et al, [24]: reported that at the 4th day of incubation on the inner side of kidney tube, primitive genital ridge had already appeared and began to depart from kidney, but had no obvious boundaries and shared the same parenchyma with kidney tube, which suggested that some kidney cells had involved in the formation of genital ridge. Here, PGCs had already migrated to the primitive genital ridge and were nearly arranged in epithelial thickening zone of body cavity which seemed like a slight belt The primordial germ cells were identified by very large size as compared to neighboring somatic coelomic epithelial cells. These primordial germ cells were large, rounded, oval or irregular in outline. Their nuclei were large vesicular, eccentric and contained one or two distinct nucleoli. Their cytoplasm was faintly stained and contained vacuoles of different shape and size. Quail primordial germ cells showed PAS negative reaction, their number was approximately equal in the left and right genital ridges at these early stages of embryonic development. Similar finding were recorded by [25, 28, 32, 33, 34, 42,53, 62, 68, 84, 88, 111].Primordial germ cells (PGCs) were identified by their remarkably large size, large spherical nuclei and refractive cytoplasmic lipids, coupled with a histochemical marker such as periodic acid-Schiff (PAS) which stains glycogen, or immunohistochemical staining of cell surface antigens such as embryonic mouse antigen-1 (EMA-1) and stage-specific embryonic antigen-1 (SSEA-1) [32, 33, 34, 53, 62, 68, 109] but Clara et al., [25] found no glycogen granules in the quail PGCs cytoplasm at any stage of embryonic period. The present result indicated that primordial germ cells were originated extragonadal, where these cells could be seen migrating in the dorsal mesentery of the hind gut to rd the presumptive gonads and circulating within the intra embryonic blood vessels near the genital ride at this early stage of embryonic development. Similar findings were demonstrated by several studies [28, 42, 45, 58, 63, 93, 95].Intrapat (2011). D’costa et al., [28] stated that during day 2 and 3 equal number of PGCs were colonized the right and left gonads, a higher number of PGCs were observed at day 4.Explanation of migration of germ cell in avian embryos were described by several authors: According to Ukeshima et al., [95] PGCs in circulation extend filopodia that come in contact with the inner surface of the endothelial cell; they adhere to the blood vessels and move through the gaps between the endothelial cells by dia pedesis. PGCs at (stage 20 at Hamburger-Hamilton (HH) and onwards actively migrate through the dorsal mesentery, enter into the germinal epithelium and colonize the gonad [40, 58]. Germ cells moving through the mesenchyme were often amoeboid in shape and contained microvilli and cytoplasmic processes [93]. Nakamura et al., [68] clarified that unlike other species, in avian embryos PGCs use the developing vascular system as a vehicle for transport from the germinal crescent region to the future gonadal region. While Intrapat, [45] mentioned that at the beginning of chicken PGCs migration toward the presumptive gonad they migrated passively, the germinal crescent PGCs migrated through endothelial cells of dorsal aorta via “diapedesis”. This step has been called passive migration which was a type of ameboid movement in vivo, occurring in blood chicken PGCs which had pseudopodia. Intrapat, [45] described there are two different sizes of primordial germ cells (PGCs) circulating and tissue types. PGCs have been shown to use extra embryonic blood vessels as the route to circulate throughout the entire embryo; therefore, these cells have been called “circulating-PGCs” (cPGCs). The morphology of circulating-PGCs has been described as round and with protruding cytoplasmic processes inserting between endothelial cells to migrate out of the embryonic blood vessels. The circulating-PGCs that have left the blood circulation incorporated into and migrated along mesenchymal cells in the dorsal menstery at the level of the mesonephros. Hence, those PGCs have been called “tissue-PGCs” (tPGCs). The morphology of PGCs at this step was different from c PGCs since they display pseudopodia, which were also characteristic of cells undergoing amoeboid movement. Hence, the process that allowed tissue-PGCs to migrate actively along migratory routes by themselves has been called “active migration.” He added that PGCs stop active migration when they reach the genital ridges. Where the genital ridge formation was related to the implantation of migrating PGCs. PGCs which have implanted in the genital ridges have been called “gonadal-PGCs (gPGCs)”. Furthermore, it has been shown that vascularization at the level of the genital ridges was crucial for PGC implantation in the genital ridges since the lack of this process has been implicated in PGC migration in chicken embryos. Chick germ cells originated in an extraembryonic area during the early stages of development, and then migrated into the forming right and left gonadal analogues via blood circulation [93] .After the arrival of the germ cells, the gonadal primordium begins to differentiate into either an ovary or a testis according to the genetic sex. In female chicks, however, the ovary was developed only on the left side, and the right one degenerated. The present work revealed that as development proceeded and at 5 days quail embryo the indifferent gonads were increased in size and became bulged into the coelomic cavity as distinct organs from the mesonephros. The left gonad was slightly larger than the right one, but it was not possible to determine from its histology or morphology whether it will become a testis or an ovary, and this was the indifferent stage. These gonads were formed of mass of epithelial cell cords covered with outer surface epithelium and these cords were appeared as a bud like extension from the surface epithelium, similar to findings were demonstrated by [26, 31, 37, 48, 63,71, 79, 81, 83]. According to the opinion of different authors for a description of gonads formation, there are different descriptions. Romanoff [79] mentioned that on the fifth day of chick incubation period, the sex cords of the first proliferation were formed as a buds from the germinal epithelium and grow into the deeper lying stroma, these cords were elongated, anastomosed and carried primordial germ cells with them, during this indifferent stage there was no morphological distinction between the sexes. Franchi et al, [31] Nieuwkoop and Sutasurya, [71] recorded that proliferation of the surface epithelium of the indifferent gonad invades the underlying mesenchyme. These cells gave rise to sex cords; these cords did not branch, but elongated and became separated from the cortical region of the gonad by primary tunica albuginea. However, Witschi [105, 108] reported that in birds theses cords were derived from the mesenchymal blastema of the indifferent gonads. Blocker [15] in his studies on the English sparrow came to the same conclusion. On the other hand, some authors suggested that at the 5th day of incubation the gonads began to differentiate into testis or ovaries [24, 70, 80]. The present investigation showed different stages of mitotic division in the primordial germ cells and increase in their in number at the early developmental stages, this result was similar to the finding of [88, 106] that these cells divided during migration. However, Franchi et al. [31] and Zuckerman and Baker [111] stated that primordial germ cells undergo a period of mitotic proliferation once they have reached the genital ridge and that, in general, those cells that failed to reach their final distribution were degenerated and disappeared. The present study showed that at the seventh day of incubation the ovaries were distinguished morphologically and histologically, so sex differentiation could be demonstrated at this stage. Morphologically, the left ovary was larger and somewhat dorsal to the right one, moreover, the left ovary was located somewhat cranial to the right. They were converged caudally to become in contact with each other at their caudal end and similar to [28, 48, 50, 59, 70, 71, 79, 88, 85]. According to Li et al. [59] the gonadal differentiation of the chicken embryo began on the 6th day of the incubation period and this kind of differentiation became more significant on the 7th day. Ottige [72] reported that the chick gonads sex differentiation could be demonstrated during the first half of incubation, whereas the hypothalamic – hypophyseal-gonadal axis become functional. Histologically the left ovary could be identified by the organization of the ovary into the primitive outer cortex and inner medulla. The cortex was formed of cortical cords that extend inward from the gonadal epithelium, while the medulla was composed of medullary cords and the enlarged inner spaces (medullary lacuna). During development the thickness of the cortex was increased as a result of proliferation of both germ cells and somatic cells, also the medulla was increased as a result of formation of additional medullary cords and distensions of the others. Similar findings were obtained by [28, 36, 50, 79, 88, 90, 96, 97, 98]. Several authors described the increased thickness of cortex by different explanation.D’costa et al [28] reported that by the sixth and seventh day of incubation, the primary sex cords in the medullary region of the left ovary were break up and the PGCs were embedded into the secondary set of cords forming the outer cortical region. The cortical region formed the major component of the left ovary. Romanoff [79] mentioned that the sexual differentiation happened on the seventh day and the sexual manifestation of the female gonads was different in the right and left ovaries. Whereas the right ovary was undergone degeneration, the left one was increased in size. The left ovary was composed of cortex and medulla, the ovarian cortex (secondary cords) formed by inward growth of the surface epithelium. The majority of germ cells were included and proliferated in this region to form germ cells clusters. Also, some germ cells were found in the medulla [97]. Kingston and Bumstead [56] suggested that on the 8th day of hatching, the PGCs in the ovary began to differentiate into oogonia. The present study showed that oogonia were found on the 7th day which demonstrated that the differentiation of quail’s PGCs was earlier than the chicken’s that led to the time difference of the early gonadal development The results showed that the disparity in size between the left and right ovaries was well marked at all examined ages; the right ovary was much smaller than the left ovary. Similar results were obtained by [37, 38, 46, 50, 80,99]. Some authors proposed that these differences between left and right ovaries depend on the asymmetric gene expression between them, which occurred only in the cortex before sex differentiation. Asymmetric PITX2 gene expression in the left presumptive gonad leads to the asymmetric expression of the retinoic acid (RA)-synthesizing enzyme, RALDH2, in the right presumptive gonad. Subsequently, RA suppresses expression of the nuclear receptor Ad4BP/SF-1, cyclin D1, and estrogen receptor an in the right presumptive gonad. As a consequence, inhibition of cell proliferation and loss of responsiveness to estrogen in the right ovarian primordium are induced [37, 50, 78] In mammals, the gonads develop bilaterally through the orchestrated action of a number of genes [50, 82, 87]. Unlike mammals, most female birds developed ovary only on the left side [50]. Guioli et al [28] studied the gonadal a symmetry and sex determination in birds, they reported that in chick embryos as in mammalian embryos, the gonads raised as a thickening of the coelomic epithelium which formed a ridge running in an anterior: posterior orientation on the ventral surface of each mesonephros (primitive kidney). Gonad genesis began at around 72 h of development, but the genital ridges were not macroscopically evident. As development proceeded, the morphological appearance of the gonads was initially similar in males and females. Throughout embryonic development, male and female gonads continued to increase in size and immediately prior to hatching, the right gonad was around 2.6 mm by 0.5 mm and the left which is clearly an ovary is 8 mm by 1.5 mm. Also, they mentioned that in the initial stages of gonad genesis, the epithelial ridge was composed of columnar cells that overlie tissue that lacks the typical nephron structure characteristic of the underlying mesonephros and that was instead composed of clusters of epithelial-like cells embedded in mesenchyme. The ‘genital ridges’ protrude into the coelomic cavity as distinct organs and comprise a pseudostratified the columnar epithelium (germinal epithelium covering a central core (medulla) organized into epithelial cords. The origin of these so-called primitive sex cords, which were also apparent in some mammals such as humans, but not the mouse,was still unclear. There is also a clear Left: Right asymmetry in the distribution of the germ cells. The left gonad contains more germ cells than the right. This asymmetry has been observed in the chick and showed in other birds as quail and duck. They added that PGC colonization of the gonads was driven by chemoattractants secreted, by the gonadal mesoderm, so it was possible that the unequal L: R distribution of germ cells was simply due to an L: R asymmetry in the level of gonadal chemokines. The present study showed that the medullary germ cells in both left and right ovaries did not show meiotic division and these germ cells were exhibited sometimes a chromatin con densation similar to those of meiosis, this feature, however, did not increase the number with the advancement of development. In contrast, dead cells were coincidentally seen in the medulla and such features were never seen in the cortex. From this observation, the condensation of chromatin seemed to be an initial sign of cell death. In addition to dead germ cells were mainly found attached to the epithelia of the lacuna and sometimes germ cells were released into the lacuna as a solitary cell or as masses formed of several cells. In the right ovary, germ cells released into the lacuna were found at the holes of the ovarian surface, which were continuous with the medullary lacuna. Cortical germ cells were never released from the ovarian surface. Similar to the observation obtained by[95, 97, 98]. Release of germ cells from the ovarian surface has been observed in some mammalian species [60, 67].Motta and Makabe [67] observed the extrusion of germ cells from the ovarian surface of humans, their observation revealed that the germ cells, regarded as cortical germ cells, were directly extruded from the ovarian surface epithelium of the ovary. These observations indicated the differences in the mechanism of germ cell release between animal species. Scanning electron microscopic observation of the quail ovary in late embryonic stage (11-17 day of incubation) revealed that the normally developing left ovary was spindle-shaped and its surface appeared rough at low magnification. At higher magnification, the epithelial cells were usually round or oval cells, but sometime were fusiform in shape. The surface of each cell was bulging and had many protrusions such as microvilli or globular projections were present on the cell surface. No stomata were detected on the outer surface of the left ovary – similar results were obtained by [98]. The present study showed that the surface epithelial cells of both left and right ovaries possessed a coat with a light reaction to both PAS and Alcian blue stain and with the advancement of development, the reaction was stronger in the left ovary during development as the mucous substance increased in the ovary similar to findings of Avila et al [8]. Avila et al [8] reported that both germ cells and epithelial cells possessed a coat with a light reaction in the right ovary and more intense reaction in the left one when they stained with PAS, Alcian blue or Ruthenium red, the mucous substance were increased from 15th day. They mentioned that mucous substance may play a role in cellular migration in the left ovary, this not in the right ovary which suffered from regression and embryonic atrophy. Quantification of the germ cells in quail ovary in different stages of development revealed that the germ cell population per unit area was few at an early stage of embryonic develop- ment. The number of germ cell then gradually increased and reached a maximal value at 15 days of incubation with the advancement of the embryonic development, the germ cells declined gradually and reached a minimal value at 1 day after hatching. The present study showed that in quail only 70% of the peak number of germ cells was survived in the left ovary of the quail similar to the result of [43, 51, ] .Huges [43] reported that the number of germ cells in the left ovary of chicken embryo was reached the peak at stage 43 (17 days of incubation). Subsequently, the population of germ cells was declined to two-thirds of the highest number at 1 day after hatching. The decline of germ cells before hatching was due to a high incidence of pro grammed cell death (apoptosis) of germ cells. Jonshon [51] mentioned that the number of germ cells in chick embryo was increased from approximately, 28,000 on the 9th day of development to 680.000 on the 17 days, and subsequently decreases to. 480.000 by the time of hatching, when oogenesis was terminated. The germ cell loss in quail ovary was due to a phenomenon of apoptosis. It is well known that apoptosis is a type of physiological cell death that occur in proliferating and differentiating tissue occurred. The rate of germ cell loss varied considerably in different species. In human and cow, only 5% of the peak number of germ cell survived in the gonad at birth [10, 29] .13% survived in monkey and guinea pig [11. 30% removed in the newborn rat [14] while in the pig half of the germ cells survive, 20% of the peak camel fetus [1].The present study showed that in quail only 70% of the peak number of germ cells was survived in the left ovary of the quail. This considered a high rate compared with the other different species. The first evidence of the follicle formation in the present study was demonstrated in the cortical zone of the quail left ovary at one day after hatching.The transformations of oocytes cell nests defined as folliculogenesis [73]. So folliculogenesis in the left ovary of quail occurred at one day after hatching similar to findings of Kannenkeril and Domm [52].The time at which primordial follicle was initiated varied in different species. In some species, primordial follicle started to form in embryonic life as in human, monkey, horses, cow and she-camel [1, 3, 64, 57]. In some other species as in cat, dog, rabbit and birds, the primordial follicle started to form during the post-natal of development or during post-hatching period [2, 4, 43, 75]. The role which the ovarian surface epithelium played in the contribution to granulosa cell differentiation varied widely between species. In some species, the ovarian surface epithelium seemed to be negligible [20]. While in other species many cells form the outer surface epithelium entered the ovarian cortex and became associated with oocytes to form primor dial follicle [73]. The present study showed that with the advancement of embryonic development, the cortical invigilation was increased in both depth and number at the eleventh, thirteenth and seventeenth day of quail embryo incubation similar findings were obtained by Soliman [88] The ovarian surface epithelium appeared to extend into the ovarian cortex and became in contact with germ cells. This observation of the present study suggested that the ovarian surface epithelium was involved in the granulosa cell differentiation and their outer surface epithelium contributed cells to the follicle envelope [2, 83]. Hutt et al. [44] supported the hypothesis that the ovarian surface epithelium was the primary source of pre-granulosa Cells for follicle formation in the rabbit. The results of the present study clearly showed that the onset of the first meiotic prophase in the female germ cells of the quail occurred at 11 days of incubation similar to findings obtained by [28, 52, 90, 95, 96]. However, Huges et al. [43] mentioned that the meiotic germ cells in chicken embryo on the 13th day of incubation. On the other hand Callebaut [22]; Smith et al., [85]; Yu et al. [110] : reported that onset of meiosis in chicken embryo was on the 15th day of incubation. The first meiotic division of the oocyte in quail was characterized by prolonged prophase. The meiotic prophase of the oocyte was subdivided into a number of stages reflected the state and arrangement of chromosomes.This stages of meiotic prophase were leptotene, zygotene, pachytene, and diplotene. Leptotene stage of the first meiotic prophase was large rounded cells with a large spherical nucleus. The chromosome at this stage was larger strinaary structure and occupied much of the nuclear space in a reticular arrangement, these nucleus was usually not visible but it occasionally distinguishable as a lightly acidophilic body at the periphery of the nuclear membrane. Bouquet formation was a characteristics feature of zygotene stages of the first meiotic prophase was commonly demonstrated in paraffin sections stained with hematoxylin and eosin. Oocyte at pachytene stage was characterized by shorter and thickener chromosomes compared with those of the other meiotic prophase. The chromosomes during pachytene stage were thickener and were evenly distributed throughout the nucleus. The stage of meiotic prophase in the quail ovary was qualitatively similar to those described for other mammalian species [2]. The present study revealed that the differentiation was more advanced at the center of the cortex than at the cortical extremities. The same was true of amphibians, where oocytes in the more central areas of the cortex co-exist with oogonia at the periphery [104]. A similar tendency has been reported for some mammals like rabbit [103]; mouse [16]; Man [10], although the differences in the stages of development attained by peripherally, as compared with centrally placed germ cells may only be slight [18, 27, 65]. The present study showed that at early stages of embryonic development the medulla of left and right ovary contained isolated interstitial cells, and with the advancement of development these cells increased in number and were present in groups or cell nest that increased with the age. Interstitial cells exhibited a foamy cytoplasm with vacuoles Similar to findings were observed by [8, 9, 36]. Avila et al, [8] reported that since 11 days of incubation. The interstitial cells were present in groups and exhibited a foamy cytoplasm and clear vacuoles of variable size. A number of cell nests increased with the ages of the embryos. Also, they found an accumulation of sudanophilic granules and an enzymatic activity in the interstitial cells that increased during the course of development. The present study showed that the ovarian stroma was contained fine reticular fibers that were increased in amount with the development and the ovarian stromal fibers were reacted positively to PAS and Alcain blue stain, this result was supported by the finding of Soliman [88] The present study showed that the development of the gonads of the quail was similar to those of chick, despite the difference in the incubation period, 16 to 17 days for the quail and 21 for the chick the degree of development of the gonads at the time of hatching was about the same. Consequently, an embryonic development requiring 21 days for the chick was completed in a shorter period in the quail. Hence, there was evidently speeding up to on development in the case of quail, for example, in the chick [ 90, 91] th author found that the PGCs appeared in the germinal epithelium by three and one-half days of incubation whereas in quail we found that this occurred as early as two and one-half to three days. Such acceleration was also observed in the proliferation of the medulla and cortical cords and other major events. Gonzalez- moran [36] stated that time of sex differentiation in chicken was the 8th day of the incubation and we found that this occurred at the 7th day of the incubation. In addition to, Huges [42] mentioned that the fist meiotic prophase in chicken was detected in the13-14th day of incubation, while the present study revealed that this occurred on the 11th day in quail.
SUMMARY
The material used for the present study consists of quail embryos(Coturnix Coturnix. Japanica) collected during 3, 5, 7, 9, 11, 13, 15, 17 days of incubation and one day after hatching. The materials obtained from research farm center faculty of agriculture Assiut University. The materials were fixed in the proper fixatives and prepared for microscopic examination. Other samples were taken for scanning electron microscope. The histomorphological studies of the development of the ovary in quail revealed that at an early stage of embryonic development (third day of incubation) the indifferent gonad could be identified as bilateral cellular thickening located ventromedial to the mesonephros. Each gonad was represented by thickened coelomic epithelium contained few number of primordial germ cells. The primordial germ cells were very large as compared to the neighboring somatic coelomic epithelial cells. The primordial germ cells were large, rounded, oval or irregular in outline. Their nuclei were large, vesicular, eccentric and contained one or two nucleoli. Their cytoplasm was faintly stained and contained vacuoles of different shapes and size. Primordial germ cells showed a negative reaction to both PAS and alcian blue stain. As development proceeded (at 5-day quail embryo), the indifferent gonads were slightly increased in size and became bulged into the coelomic cavity there was an asymmetry between the left and right gonads where the left gonads were larger than the right one. Each gonad was formed of a core of mesenchymal connective tissue contained few clusters of primordial germ cells and covered from outside by a single layer of cuboidal or pyramidal cells. The gonadal germinal epithelium showed an outer thin positive coat to PAS and alcian blue stains. Gonadal stroma showed also PAS and alcian blue stains. The reticular fibers appeared black when stained with Gomoris reticulin technique. Histological sex differentiations were demonstrated at the seventh day of incubation. The left ovary of quail could be identified by the presence of the primitive outer cortex and primitive inner medulla, the primitive cortex was separated from the inner primitive medulla by a thin layer of connective tissue. The outer surface epithelium of the left ovary was formed of a single layer of cuboidal or columnar cells in some areas whereas in other regions was multilayered. Some germ cells were interposed among surface epithelial cells and showed mitotic division. Most germ cells in the left ovarian cortex at this stage of embryonic development were oogonia The oogonia were smaller than the primordial germ cells. Their nuclei were large, vesicular and contained 1-3 distinct nucleoli. Mitotic figures were frequently demonstrated at this stage of embryonic development. Oogonia in prophase, metaphase, anaphase and telophase of the mitotic division could be demonstrated in the left ovarian cortex at this stage of development. Normal oogonia comprised the most germ cell population, while degenerated or apoptotic germ cells were still very few and rarely demonstrated. The ovarian medulla was formed of medullary cords, blood vessels, and lacunar channel system. The interstitial cells were rounded, oval, or polyhedral cells. Their nucleoli were vesicular and eccentric. Their cytoplasm was pale acidophilic. The epithelial cells of the lacunar channels of the ovarian medulla were formed of low cuboidal or flattened cells, their nuclei were rounded, vesicular and contained distinct nuclei. Free germ cells in the medullary lacunae were frequently demonstrated in both right and left ovaries. With the advancement of the development and at 11-17 day of embryo development the left growing quail ovary was more organized into an outer cortex and inner medulla. The germ cells in the left ovarian cortex were arranged in small groups or nests. Each group was surrounded by a layer of the connective tissue showed a positive reaction to PAS and alcian blue stains. The female germ cells contained two populations; the first population was oogonia in premeiotic resting phase. The second population was oocyte in first meiotic prophase. Leptotene, zygotene, pachytene and diplotene meiotic prophase stages could be demonstrated. Zygotene stage of meiotic prophase was more demonstrated than another prophase stage. Meiotic germ cells were first detected only in the left ovarian cortex of quail from the day 11 of the embryonic life. The number of germ cells was higher than the previous embryonic stage.The medulla of the left and right ovaries presented the same structure as in the previous embryonic stage. The first evidence of primordial follicle formation was demonstrated in the left ovarian cortex of quail at one day after hatching. The primordial follicle was formed of an oocyte surrounded by a single layer of flattened follicular cells. The oocytes of the primordial follicle were demonstrated at the resting diplotene stage of the first meiotic division. The primordial follicles either present singly or in groups forming follicle nest. Quantification of the germ cells in quail during different stages of embryonic development revealed that the germ cell population per unit area was few at an early stage of embryonic development. The number of germ cells was gradually increased with the advancement of embryonic development and reached the maximal value at 15th day of incubation with the advancement of development, the germ cells declined gradually and reached a minimal value at one day after hatching. The present study showed that in quail only 70% of the peak numbers of germ cells was survived in the left ovary of the quail.
CONCLUSION
The ovary in quail revealed that at an early stage of embryonic development (third day of incubation) the indifferent gonad could be identified. Morphological and Histological sex differentiations were demonstrated at the seventh day of incubation. The left ovary of quail could be identified by the presence of the primitive outer cortex and primitive inner medulla. At 9 days of embryo development, the surface of the cortex showed different degrees of invaginations that divided the cortex into numbers of a lobes-like structure With the advancement of the development and at 11-17 day of embryo development the left growing quail ovary was more organized into an outer cortex and inner medulla. Meiotic germ cells were first detected only in the left ovarian cortex of quail from the day 11 of the embryonic life. The ovarian cortex at 7 days of embryo development contained most oogonia and few primordial cells. At 11-17 day of embryo development the germ cells in the left ovarian cortex were arranged in small groups or nests and showing two populations; the first population was oogonia in premeiotic resting phase. The second population was oocyte in first meiotic prophase. The ovarian medulla was formed of medullary cords, blood vessels, and lacunar channel system. The epithelial cells of the lacunar channels of the ovarian medulla were formed of low cuboidal or flattened cells, their nuclei were rounded, vesicular and contained distinct nuclei. Free germ cells in the medullary lacunae were frequently demonstrated. The first evidence of primordial follicle formation was demonstrated in the left ovarian cortex of quail at one day after hatching. The outer surface epithelium of the left ovary was formed of a single layer of cuboidal or columnar cells in some areas whereas in other regions was multilayered.The gonadal germinal epithelium showed an outer thin positive coat to PAS and alcian blue stains.
REFERENCES
- Hafez EA. (2004). Histomorphological studies on the development of the ovary of one humped camel. Ph. D. Thesis, Fac. of Vet. Med Assuit University, Assuit, Eygpt.
- Kareem AM. (2010). Histomorphological studies on the development of the ovary of the rabbits during the postnatal period. M.V.Sci Thesis, Fac. of Vet. Med., Assuit University, Assuit, Eygpt.
- Abuzaid RMM. (1990). Some prenatal morphological studies on the ovary of the water buffalo in Egypt. Thesis, Fac. of Vet. Med. Suez Canal University, Egypt.
- Andersen AC and Simpson ME. (1973). The ovary and the reproductive cycle of the dog (Beagle). Geron-X, Inc Los Altos California.
- AL-Thani R and Simkiss K. (1991). Effect of an acute in vivo application of concanavalin A on the migration of avain primordial germ cell. Protoplasm. 161: 52-57.
- Al-Saffar FJ and Abood DA. (2014). Histomorphological study of the pre hatching development of the female genital system in Indigenous Mallard Duck (Anas platyrhynchos). International Journal of Advanced Research. 2(10) : 248-263.
- Ando Y and Fujimoto T. (1983). Ultrastructural evidence that chick primordial germ cells leave the blood-vascular system prior to migration to the gonadal anlagen. Dev. Growth Differ. 25(4): 345-352.
- Avila RE, Samar ME and de Fabro SP. (1991). Evolution of germinal epithelium in the chick ovary during embryogenesis. Rev. Fac. Cienc. Med. Univ. Nac. Cordoba. 49(1): 11-15.
- Avila RE, Samar ME and De Fabro SP. (1989). Structural, ultrastructural and cytochemical study of interstitial cells of the chick ovary during embryo development. Rev. Fac. Cienc. Med. Univ. Nac. Cordoba. 47(1-2): 9-12
- Baker TG. (1963). A quantitative and cytological study of germ cells in human ovaries. Proc. R. Soc. Lond. Ser. B. 158(972): 417-433.
- Baker TG. (1966). A quantitative and cytological study of oogenesis in rhesus monkey. J. Anat. 100: 761-776.
- Bancroft JD, CLayton and Suvarna SK. (2013). Bancroft’s Theory and Practice of Histological Techniques. ed 7. London, Churchill Livingstone.
- Baumgartner J. (1994). Japanese quail production, breeding and genetics. World�s Poult. Sci. J. 50: 227-235.
- Beaumont HM and Mandl AM. (1962). A quantitative and cytological study of oogonia and oocytes in the foetal and neonatal rat. Proc. Ray. Soc. B. 155(961): 557-579.
- Blocker HW. (1933). cited by Romanoff (1960). Text book of the Avain Embryo, Structural and Functional development. First edition, The Macmillon company, Newyork, Brett-Macmillon Ltd. 1-87.
- Borum K. (1961). Oogenesis in the mouse. A study of meiotic prophase. Exp. Cell Res. 24: 495-507.
- Brambell FWR. (1927). The development and morphology of the gonads of the mouse. Part I. The morphogenesis of the indifferent gonad and of the ovary. Proc. roy. Soc. B. 101(711): 391-409.
- Brambell FWR. (1956). Ovarian changes. In «Marshall›s physiology of reproduction» (A.S Parkers, ed.) Longmans, Green and Co, London. 1: 397-542.
- Byskov AG. (1975). The role of reta ovarii in meiosis and follicle formation in different mammalian species.J.Reprod.Fertil. 45: 201-209.
- Byskov AG, Skakkebaek NE, Stafanger G and Peters H. (1977). Influence of ovarian surface epithelium and rete ovarii on follicle formation. J Anat. 123: 77-86.
- Callebaut M. (1967). Premeiosis and premeiotic DNA synthesis in the left ovary of the female chick embryo. J. Embryol. Exp. Morphol. 18(3): 299-304.
- Callebaut M. (1976). Origin of ovarian follicle cells in birds. Experientia. 32(10): 1337-1339
- Carlon N, Piznt J and Stahl A. (1983). Mesonehric origin of the gonadal primitive medulla in chick embryo.Anat Embryol. 166(3): 399-414
- Chang GB, Chen R, Qin YR, Zang Y, et al. (2012). The development of primordial germ cells (PGCs) and testis in quail embryo, pakistan veterinary journal 32(1): 88-92.
- Clara A, Ana C, Víctor N, Jesús R, et al. (2007). Carbohydrate characterization of quail primordial germ cells during migration and gonadal differentiation. J. Anat. 210(1): 98-111.
- Clinton M and Haines LC. (2001). An overview of factors influencing sex determination and gonadal development in birds, EXS. (91): 97-115.
- Cowperthwaite MH. (1925). Observations on pre- and postpubertal oogenesis in the white rat, Mus norvegicus albinus. Amer. J. Anat. 36(1): 69-89.
- DCosta S, Pardue S and Petitte J. (2001). Comparative development of avian primordial germ cells and production of germ line chimeras. Avian and Poultry Biology Reviews. 12 (4): 151-168.
- Erickson BM. (1966). Development and radio-response of the prenatal bovin ovary. J. Reprod Fertil. 11: 97-105.
- Fargeix N, Didier E and Didier P. (1981). Early sequential development on avain gonads.An Ultra structure study using selective glycogen labeling in germ cells. Reprod Nutr Dev. 21(3): 497-496.
- Franchi L, Mandle A and Zuckerman S. (1962). Development of the ovary and the process of oogensis in the ovary.London: Academic press. 1-88.
- Fujimoto T, Ukeshima A and Kiyofuji R. (1976). The origin, migration and morphology of the primordial germ cells in the chick embryo. Anat Rec. 185(2): 139-145.
- Ginsburg M and Eyal-Giladi H. (1986). Temporal and spatial aspects of the gradual migration of primordial germ cells from the epiblast into the germinal crescent of the avian embryo. J Embryol Exp Morphol. 95: 53-71.
- Ginsburg M and Eyal-Giladi H. (1987). Primordial germ cells of the young chick blastoderm originate from the central zone of the area pellucida irrespective of the embryo-forming process. Development. 101(2): 209-219.
- Gomori. (1937). Silver impregnation of reticulum in paraffin section. Amer J path. 13(6): 993-1002.5.
- Gonzalez-Moran MG. (2011). Histological and stereological changes in growing and regressing chicken ovaries during development. Anat. Rec (Hoboken). 294(5): 893-904
- Guioli S and Lovell-Badge R. (2007). PITX2 controls asymmetric gonadal development in both sexes of the chick and can rescue the degeneration of the right ovary. Development. 134(23): 4199-4208.
- Guioli S, Nandi S, Zhao D, Burgess-Shannon J, et al. (2014). Gonadal Asymmetry and Sex Determination in Birds. Development. 8(5): 227-242
- Hamburger V and Hamilton HL. (1992). A series of normal stages in the development of the chick embryo. 1951. Dev Dyn. 195(4): 231-272.
- Harris HF. (1898). A new method of ripening hematoxylin. In: Mikroskopische Technik. Romeis (ed.), Oldenburg, München.
- Hong YH, Seo DS, Jeong DK, Choi DK, et al. (1995). Migration of the primordial germ cells and gonadformation in the early chicken embryo. Asian-Aust J Anim Sci. 8(6): 557- 562.
- Hughes CG. (1963). The population of germ cells in the developing female chick. J Embryol Exp Morphol. 11: 513- 536.
- Hutt KJ, McLaughlin EA and Holland MK. (2006). Primordial follicle activation and follicular development in the juvenile rabbit ovary. Cell Tissue Res. 326(3): 809- 822.
- Intarapat S. (2011). Isolation and characterisation of chick embryonic primordial germ cells. Ph. D. Thesis. UCL (University College London).
- Intarapat S and Stern CD. (2013). Sexually dimorphic and sex independent left-right asymmetries in chicken embryonic gonads. PloS One. 8(7): e69893.
- Intarapat S and Stern CD. (2014). Left-Right Asymmetry in Chicken Embryonic Gonads. The Journal of Poultry science [Advance Publication]. 51(4): 352-358.
- Intarapat S, Sailasuta A and Satayalai O. (2014). Anatomical and Histological Changes of Reproductive Organs in Japanese Quail (Coturnix japonica) Embryos after in ovo Exposure to Genistein. Journal of Poultry Science. 13(1): 1-13.
- Ishiguro S, Minematsu T, Naito M, Kanai Y, et al. (2009). Migratory ability of chick primordial germ cells transferred into quail embryos .Reproduction and development. 55(2): 183-186
- Ishimaru Y, Komatsu T, Kasahara M, Katoh-Fukui Y, et al. (2008). Mechanism of asymmetric ovarian development in chick embryos. Development. 135(4): 677-685
- Johnson AL. (1986). Reproduction in the female. In: Avian Physiology (Sturkie PD Ed.). 4th ed. 403-431.
- Kannankeril JV and Domm LV. (1968).Development of the gonads in the female Japanese quail. J Anatomy. 123(1): 131-146.
- Karagenc L, Cinnamon Y, Ginsburg M and Petitte JN. (1996). Origin of primordial germ cells in the prestreak chick embryo. Dev Genet. 19(4): 290-301.
- Karnovsky MJ. (1965). A formaldehyde-glutaraldehyde fixative of high osmolarity for use in electron microscopy. Cell Biol. 27(2): 1A-149A.
- Kheirabadi M, Nabipour A, Rassouli MB and Dehghani H. (2015). Morphological development of ovaries in ostrich (Struthio camelus) embryo. Comp Clin Pathol. 24: 1185- 1191
- Kingston R and Bumstead N. (1995). Monoclonal antibodies identifying chick gonadal cells. Br Poultry Sci. 36: 187-195.
- Knospe C and Budras KD. (1992). Zur prenatalen entwicklung des pferdeovars. Anat Histol Embryol. 21: 306-313.
- Kuwana T, Maeda-Suga H and Fujimoto T. (1986). Attraction of chick primordial germ cells by gonadal anlage in vitro. Anat Rec. 215(4): 403-406.
- Li BC, Zhou GY, Sun SY, Qin J, et al. (2005). Study on the relationship between spermatogonium of the early chicken embryos and the development of the testis. J Animal Sci.Vet Med. 36 (7): 680-685.
- Mackacy S, Smith RA and Haig T. (1992). An investigation of the migratory potential of mouse oocytes in vitro.J Anat. 181: 437-446.
- McManus JFA. (1946). Histological demonstration of mucin after periodic acid. Nature. 158: 202.
- Meyer DB. (1960). Application of the periodic-Schiff technique to whole chick embryos. Stain Technol. 35(2): 83-89.
- Meyer DB. (1964). The migration of primordial germ cells in the chick embryo. Dev Biol. 10(1): 154 -190.
- Minh H, Smadja A, Herve JP and Orcel L. (1989). Histologic study of the gonads in ovarian differentiation during organogenesis. Arch Anat Cytol Pathol. 37(5-6): 201-207.
- Mintz B. (1959). Continuity of the female germ cell line from embryo to adult. Arch Anat micr Morph 48: 155- 172.
- Mizutani M. (2003). The Japanese quail, Laboratory Animal Research Station, Nippon Institute for Biological Science, Kobuchizawa, Yamanashi, Japan, 408-0041.
- Motta PM and Makabe S. (1986). Germ cells in the ovarian surface during fetal development in human. A three dimensional micro anatomical study by scanning and transmission electron microscopy.J submicrosco Cytol. 18(2): 271-290.
- Nakamura M, Maeda H and Fujimoto T. (1991). Behavior of Chick Primordial Germ Cells Injected into the Blood Stream of Quail Embryos. Okajimas Folia Anat Jpn. 67(6): 473-478.
- Nelsen OE. (1953). Text book of comparative embryology of vertebrates. The Blackstone company Inc.Toronto.
- Nechita EL, Todireanu C, Cotea CV and Solcan C. (2012). Histological aspects of the ovary in quail’s (coturnix coturnix japonica) embryo embryo Lucrri tiin ifice-ificevol.ifice seria Medicin Veterinar. 161-165.
- Nieuwkoop PD and Sutasurya LA. (1979). Primordial germ cells in the chordates. Cambridge: Cambridge University Press.
- Ottiger MA. (1989). Sexual differentiation in neuroendocrine system and behavior. Poult.Sci. 68(7): 979-989
- Peters H and McNatty KP. (1980). The ovary: A correlation of structure and function in mammals. Granada Publishing in Paul Elek. Ltd. 56(2)
- Ramteke J, Charde P, Zade S and Gabhane R. (2013). Comprehensive Study of Organogenesis During Embryonic Development of Japanese Quail.Int. J. of Life Sciences. 1(3): 193-197
- Ricken AM and Viebahn C. (2002). Stage-Specific Expression of the Mitochondrial Germ Cell Epitope PG2 during Postnatal Differentiation of Rabbit Germ Cells. Biology of Reproduction. 67(1): 196-203.
- Rodler D and Sinowatz F. (2011). Immunohistochemical and ultrastructural characterization of the ovarian surface epithelium of Japanese quail (Coturnix japonica). Ani. Sci. J. 82(2): 307-313.
- Rodler D, Stein K and Korbel R. (2015). Observations on the right ovary of birds of prey: a histological and immunohistochemical study. Anat Histol Embryol. 44(3): 168-177.
- Rodriguez-Leon J, Rodriguez Esteban C, Marti M, Santiago-Josefat B. et al. (2008). Pitx2 regulates gonad morphogenesis. Proceedings of the National Academy of Sciences of the United States of America. 105(32): 11242- 11247.
- Romanoff AL. (1960). Text book of the Avain Embryo, Structural and Functional development. First edition, The Macmillan company, Newyork, Macmillan Ltd. 1-87.
- Rong C, Guobin C, Yurong Q, Bichun L, et al. (2011). The development of ovary in quail’s embryo. African Journal of Biotechnology Vol. 10 (4): 712-717.
- Rosenthal N and Ellis H. (2010). High-throughput applicable genomic sex typing of chicken by TaqMan real-time quantitative polymerase chain reaction. Poultry Science.89(7): 1451-1456.
- Ross AJ and Capel B. (2005). Signaling at the crossroads of gonad development. Trends Endocrinol Metab. 16(1): 19- 25.
- Sawyer HR, Smith P, Heath DA, Juengel JL. et al. (2002). Formation of ovarian follicles during fetal development in sheep. Biol. Reprod. 66(4): 1134-1150.
- Singh RP and Meyer DB. (1967). Primordial germ cells in blood smears from chick embryos. Science, 156(3781): 1503-1504
- Smith CA, Roeszler KN, Bowles J, Koopman P, et al. (2008). Onset of meiosis in the chicken embryo; evidence of a role for retinoic acid. BMC Developmental Biology. 8: 85
- Steedman HF. (1950). Alcian blue 8GS: a new stain for mucin. Q J Microsc Sci. 914: 477-479..
- SolimanSM. (2007). prenatal development of the ovary in quail.Master Thesis, Fac. vet. Med.Menofia university sadaat city btanch, Egypt.
- Swain A and Lovell-Badge R. (1999). Mammalian sex determination: a molecular drama. Genes Dev. 13:755-767
- Swift CH. (1914).Origin and history of primordial germ cells in chick.Amer.J.Anat.15(4): 483-516.
- Swift CH. (1915). Origin of the definitive sex cells in the female chick and their relation to the primordial germ cells. Amer JAnat. 18(3): 441-470.
- Todireanu C, Cotea CV, Solcan C and Nechita L. (2012) Organogenesis of the ovary in the chick embryo after sexual differentiation.
- Thear K. (1998). Keeping quail (a guide to domestic and commercial management), third edition, publishedby Broad Leys publishing company London E9 5EN
- Ukeshima A and Fujimoto T. (1978). Scanning Electron Microscopy of Primordial Germ Cells in Early Chick Embryos. J. Electron Microsc. 27(1): 19-24
- Ukeshima A and Fujimoto T. (1984). Ultrastructure of primordial germ cells in the early chick embryo. In: Ultrastructure of Reproduction, eds. J. Van Blerkon and P.M. Motta. Martinus Nijhoff Publishers, Boston. 12- 18
- Ukeshima A, Kudo M and Fujimoto T. (1987). Relationship between genital ridge formation and settlement site of primordial germ cells in chick embryos. Anat. Rec. 219(3): 311-314.
- Ukeshima A and Fujimoto T. (1991) A fine morphological study of germ cells in asymmetrically developing right and left ovaries of the chick. Anat. Rec. 230(3): 378-86.
- Ukeshima A, Yoshinaga K and Fujimoto T. (1991). Scanning and transmission electron microscopic observations of chick primordial germ cells with special reference to the extravasation in their migration course. J. Electron Microsc.Tokyo. 40(2): 124-128.
- Ukeshima A. (1994). Abandonment of germ cells in the embryonic chick ovary: TEM and SEM studies. Anat Rec. 240(2): 261-266.
- Ukeshima A. (1995). Surface Morphology of quail ovaries in late embryonic stages. Okajimas Folia Anat. Jpn. 72(5): 249-258.
- Ukeshima A. (1996). Germ cell death in the degenerating right ovary of the chick embryo. Zoolog. Sci., 13(4): 559- 563.
- Vali N. (2008). The Japanese quail. International Journal of Poultry Science. 7 (9): 925-931.
- Wang Q, Weng H, Chen Y, Wang C. et al. (2015). Early development of gonads in Muscovy duck embryos Br Poult Sci. 56(4): 390-397.
- Weigert C. (1898). Über eine Methode zur Färbung elastischer Fasern. ZL Pathol 9: 289-292.
- Winiwarter HDE. (1901). Recherches sur l’ovogenese et l’organogenese de l’ovaire des mammiferes (lapin et homme). Arch. Biol. Paris et Liege. 17: 33-199.
- Witschi E. (1914). Experimentelle Untersuchung iiber die Entwicklungsgeschichte der Keimdriisen von Rana temporaria. Arch mikr Anat. 85: 9-113.
- Witschi E. (1935). Origin of asymmetry in the reproductive system of birds. American journal of anatomy. 56(1): 119-141.
- Witschi E. (1948). Migration of the germ cells of human embryos from the yolk sac to the primitive gonadal folds. contrib. Embryol.Carnegie Inst. 32: 67-80.
- Witschi, E. (1956). Development of vertebrates. W.b. Saunders co. Philadelphia.
- Witschi E. (1960). Sex and secondary sexual characters.Biology and comparative physiology of birds. Ed. A.J.Marshall, Academic Press. NewYork and london. 2: 115-168.
- Yoshinaga, K, Nakamura M and Ukeshima A. (1993). Ultrastructural characteristics of primordial germ cells in the quail embryo. Anat. Rec. 236(3): 547-552.
- Yu M, Yu P, Leghari IH, Ge C. et al.(2013). RALDH2, the enzyme for retinoic acid synthesis, mediates meiosis initiation in germ cells of the female embryonic chickens. Amino Acids. 44(2): 405-412.
- Zuckerman and Baker Jw. (1977). Text book of the ovary Vol. II Academic Press. INC. (London).