Previous Issues Volume 1, Issue 1 - 2016
Antitumor Acetylenic Lipids
Anna Kilimnik1 , Dmitry V Kuklev2 and Valery M Dembitsky3
1National Institute of Water and Atmospheric Research, Private Bag 14-901, Wellington 6241, New Zealand.
2Department of Biological Chemistry, University of Michigan Medical School, Ann Arbor, MI 48109, USA.
3Institute for Drug Discovery, P.O. Box 45289, Jerusalem 91451, Israel.
Corresponding Author:Valery M. Dembitsky, Institute for Drug Discovery, 8 Ha-Marpe Str, P.O. Box 45289, Jerusalem 91451, Israel,
Tel: +972-526877444; E-Mail: [email protected]
Received Date: 14 Mar 2016
Accepted Date: 21 Mar 2016
Published Date: 23 Mar 2016
Copyright © 2016 Dembitsky VM
Citation: Kilimnik A, Kuklev DV and Dembitsky VM. (2016). Antitumor Acetylenic Lipids. Mathews J Pharm Sci. 1(1): 005.
ABSTRACT:
This article describes antitumor acetylenic lipids and related compounds obtained from living organisms. Acetylenic lipids belong to a class of molecules containing triple bond(s). They are found in algae, plants, fungi, microorganisms, and marine invertebrates. Although polyacetylenes are common as components of terrestrial plants, fungi, and bacteria, it is only within the last 50 years that biologically active polyacetylenes having unusual structural features have been reported from plants, cyanobacteria, algae, invertebrates, and other sources. Naturally occurring aquatic acetylenes are of particular interest since many of them display important biological activities and possess antitumor, antibacterial, antimicrobial, anti-fungal, HIV inhibitory and immunosuppressive properties. There is no doubt that they are of great interest, especially for the medical, pharmacological, medicinal chemistry, and/or pharmaceutical industries. This review presents structures and describes cytotoxic activities of more than 90 acetylenic lipids, including fatty acids, glycerides, sterols and carotenoids isolated from living organisms.
KEYWORDS:
Acetylenic; Polyacetylenes, Antitumor; Sterols; Carotenoids; Fatty Acids; Lipids.
INTRODUCTION:
Natural acetylenic and/or polyacetylenic metabolites have been isolated from a wide variety of plants, fungal species, and marine algae and invertebrates [1-10]. Many of them display important biological activities, namely antitumor, antibacterial, antimicrobial, antifungal, and other chemical and medicinal properties [11-14]. More than 1000 acetylenic metabolites have been isolated and identified from plants, fungi, micro-organisms, and other organisms [1-5,11,12,15-20].
Naturally occurring acetylenic lipids possessing an acetylenic unit, as well as polyacetylenes, are of particular interest as many of them display important biological activities, namely antitumor, antibacterial, antimicrobial, antifungal, and others. This paper describes acetylenic antitumor lipids that are deemed as naturally occurring.
Acetylenic Fatty Acids and Glycerides:
The antitumor acetylenic compounds 2-cis-dehydro-matricaria acid (1), 2-trans-dehydromatricaria acid (2), cis-dehydromatricaria Me ester (3), and trans-dehydromatricaria Me ester (4) were obtained from roots of Solidago virga-aurea (Compositae) [21,22], and also from other species [23-26].
Polyacetylene (-)-17-hydroxy-9,11,13,15-octadeca-tetraynoic acid (5), referred to as minquartynoic acid, from M. guianensis stem bark showed cytotoxic activity against P-388 murine lymphocytic leukemia in vitro. The P-388 ED50 of the pure compound was 0.2 µg/mL, and it was also active in the brine shrimp larvicidal bioassay with an LC50 of 5 µg/mL.
These biological activities could account for the alleged efficacy of the plant in folk usage [27,28]. More recently, minquartynoic acid was isolated from the air-dried bark of Coula edulis, the twigs of Ochanostachys amentacea (both plants belonging to the Olacaceae), and from Minquartia guianensis bark [29-32]. In addition, acetylenic acids (6 and 7) were isolated from the twigs of Ochanostachys amentacea [31,32], and their cytotoxic activities determined (Table 1).
Minquartynoic acid (5) also showed moderate in vitro activity against Plasmodium falciparum and Leishmania major, and strongly inhibited phytohaemagglutinin A - induced proliferation of human lymphocytes [33].
Three acetylenic acids: octadeca-8,10-diynoic (8), (Z)-octadec- 12-ene-8,10-diynoic (9), and octadeca-8,10,12-triynoic (10) acids were isolated from aqueous acetone extracts of stems and leaves. All compounds showed inhibitory activity against cancer cell invasion (MM1) in vitro (Table 2) [34,35]. Five C16-acetylenic fatty acids, hexadec-8-ynoic acid (11), hexadec-10-ynoic acid (12), hexadeca-8,10- diynoic (13), hexadeca-6,8,10-trienoic (14), and hexadeca- 8,10,12-triynoic (15) acids were prepared and their inhibitory activity against cancer cell invasion examined [36].
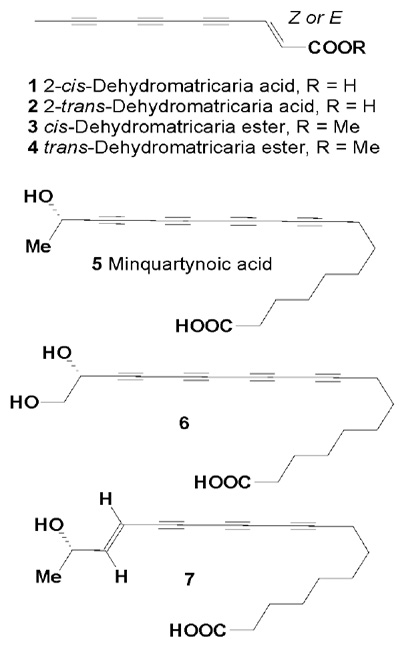 Table 1: Cytotoxic activity for compounds (1-7) (ED50,g/mL).
Table 1: Cytotoxic activity for compounds (1-7) (ED50,g/mL).
|
Cell lines |
5 |
6 |
7 |
|---|---|---|---|
|
BC1 |
3.5 |
> 20 |
> 20 |
|
Lu1 |
4.1 |
14.6 |
> 20 |
|
Col2 |
5.5 |
> 20 |
9.9 |
|
KB |
3.5 |
> 20 |
> 20 |
|
BC1 |
3.7 |
> 20 |
> 20 |
|
BC1 |
3.5 |
>2.6 |
1.1 |
|
KB-V+ |
2.8 |
11.9 |
> 20 |
|
KB-V- |
4.3 |
13.4 |
> 20 |
|
LNCaP |
1.6 |
9.2 |
0.30 |
|
SW626 |
4.1 |
19.4 |
0.36 |
|
KSKNSH |
1.4 |
6.7 |
3.7 |
|
M109 |
3.7 |
10.1 |
5.4 |
Key to cell lines used: BC1, human breast cancer; Lu1, human lung cancer; Col2, human colon cancer; KB, human oral epidermoid carcinoma; KB-V+, multidrug-resistant KB assessed in the presence of vinblastine (1 Corresponding AuthorCorresponding Author µg/mL); KB-V-, multidrug-resistant KB assessed in the absence of vinblastine; LNCaP, hormone-dependent human prostate cancer; SW626, human ovarian cancer; SKNSH, human neuroblastoma cancer; M109, mouse lung cancer.
The results indicated that the origin C18-triyne fatty acid (10) and the synthetic C16-triyne fatty acids (14 and 15) inhibit cancer cell invasion in spite of their simple chemical structures.
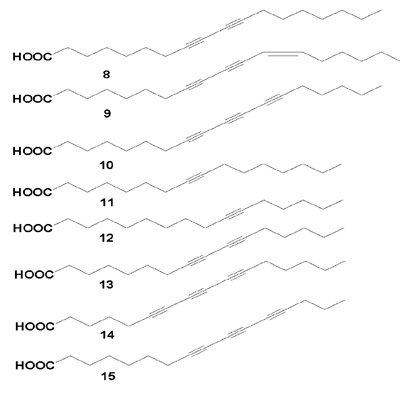
Three acetylenic metabolites (16-18) were isolated from the sponge Stelletta sp. collected from Gagu-Do, Korea [37]. These compounds exhibited no significant antimicrobial activity and displayed only weak cytotoxicity against the human leukemia cell-line K562 with LC50 values of 43, 51, and 62 µg/mL for (16- 18), respectively.
The acetylenic acid (19) and its methyl ester (20) were isolated from the sponge Xestospongia muta. The EDs of 19 for 50% inhibition in vivo PS and L1210 cell culture evaluations were 24 and 34 µg/mL, respectively, and the corresponding doses of 20 were 29 and 34 µg/mL [38]. Similar brominated fatty acids (21 and 22) were isolated from an Indonesian sponge, Oceanapia sp. [39]. Their common structural feature is a (13E, 15Z)-14,16-dibromodiene terminus. Both compounds are unstable oils. The mixture exhibits mild cytotoxicity towards KB cells. A methanol-soluble extract of the frozen marine sponge Petrosia sp. showed significant activity in the brine shrimp larvae lethality bioassay (LD50 30 µg/mL), and showed cytotoxic activities against a panel of human solid-tumor cells.
Twelve brominated acetylenic acids, (23a-d, 24a-d, and 25), together with the known compound (26) were isolated from marine sponge Xestospongia testudinaria (Nepheliospongiidae) [40]. All isolated brominated acetylenic acids showed weak cytotoxicity against the tumor cell lines HeLa, and HL-60. Pellynic acid (27) inhibited inosine monophosphate dehydrogenase with an IC50 of 1 pM/mL [41].
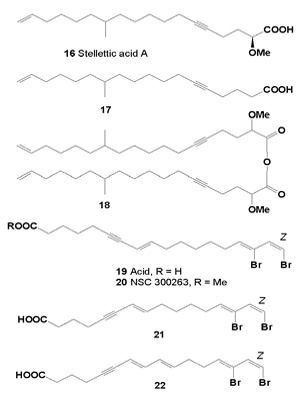 Table2Inhibitory activities of C16 and C18-acetylenic acids on cancer cell invasion (MM1).
Table2Inhibitory activities of C16 and C18-acetylenic acids on cancer cell invasion (MM1).
|
Compound |
Concentration (µg/mL) |
Inhibitory activity (%) |
|---|---|---|
|
8 |
10 |
61.1 |
|
9 |
10 |
89.8 |
|
10 |
10 |
99.4 |
|
10 |
5 |
94.9 |
|
10 |
2.5 |
45.6 |
|
11 |
10 |
82.4 |
|
12 |
10 |
77.2 |
|
13 |
10 |
85.6 |
|
14 |
10 |
95.7 |
|
14 |
5 |
85.4 |
|
14 |
2.5 |
50.3 |
|
15 |
10 |
98.7 |
|
15 |
5 |
90.7 |
|
15 |
2.5 |
60.5 |
Montiporic acids A and B were not only antibacterial against Escherichia coli, but also cytotoxic activities against P-388 murine leukemia cells, with IC50 values of 5 and 12 µg/mL, respectively. Cytotoxic activity of compounds 29, 30, 32 and 33 showed in Table 3.
Many species of tunicata produce bioactive compounds [42]. Callysponginol sulfate A (35), a sulfated C24 acetylenic fatty acid from the marine sponge Callyspongia truncata, is a membrane type 1 matrix metalloproteinase (MT1-MMP) inhibitor. Compound 35 inhibited MT1-MMP with an IC50 value of 15 µg/mL, and sodium 1-(12-hydroxy)-octadecanyl sulfate was isolated from a marine tunicate as a matrix metalloproteinase 2 (MMP2) inhibitor [43,44]. This compound inhibited MMP2 with an IC50 value of 9.0 µg/mL.
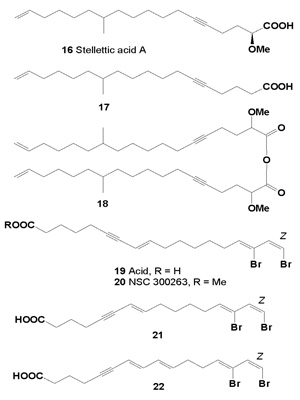 Table 3Cytotoxic activities (ED50 �g/mL) of compounds against human solid tumor cells.
Table 3Cytotoxic activities (ED50 �g/mL) of compounds against human solid tumor cells.
|
Compound |
A549 |
SK-OV-3 |
SK-MEL-2 |
XF498 |
HCT15 |
|---|---|---|---|---|---|
|
29 |
> 30 |
20.52 |
> 30 |
> 30 |
25.61 |
|
30 |
> 30 |
> 30 |
> 30 |
> 30 |
> 30 |
|
32 |
> 30 |
> 30 |
> 30 |
> 30 |
> 30 |
|
33 |
6.31 |
7.50 |
7.97 |
7.72 |
8.30 |
|
34 |
> 30 |
20.52 |
> 30 |
> 30 |
25.61 |
Key to cell lines used: A549 = human lung cancer; SK-OV-3 = human ovarian cancer; SK-MEL-2 = human skin cancer; XF498 = human CNS cancer; HCT15 = human colon cancer.
The genus Montipora (phylum Cnidaria) is very rich in acetylenic compounds and many of them were shown to be cytotoxic and/or to possess antifungal and antibacterial properties. Two polyacetylene carboxylic acids, montiporic acids A (28) and B (31), have been isolated from the eggs of the scleractinian coral M. digitata [45]. They exhibited antimicrobial activity against Escherichia coli and cytotoxicity towards P-388 murine leukemia cells (Table 3).
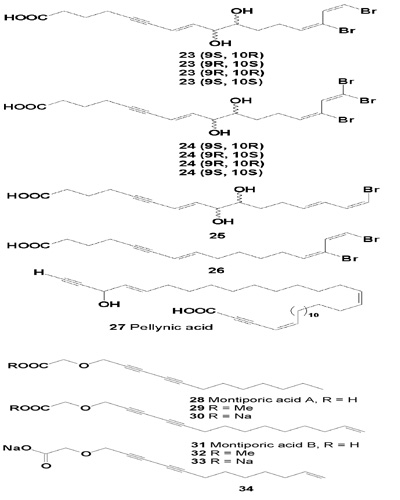
The structures of octadec-5-yne-7Z,9Z,12Z-trienoic acid (liagoric acid) (36), 7-hydroxy-octadec-5-yne-9Z,12Z-dienoic acid (37), and glyceryl octadec-5-yne-7Z,9Z,12Z-trienoate (38) isolated from Liagora farinosa are presented below. These compounds showed acute toxicity toward Eupomacentrus leucostictus at concentrations from 5 to 8 µg/mL. It was shown that at a concentration of 31 µM, liagoric acid inhibited cyclooxygenase activity [46,47].
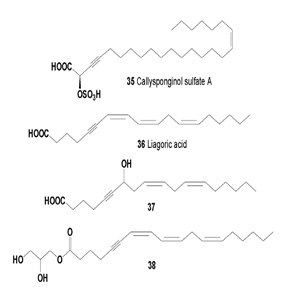
The extract obtained from the leaves of Anacolosa pervilleana was selected for its significant activity in assays. One new (E)- tridec-2-en-4-ynedioic acid named anacolosine (39), together with three known acetylenic acids, the octadeca-9,11,13-triynoic acid (40), (13E)-octadec-13-en-9,11-diynoic acid (41), (13E)-octadec-13-en-11-ynoic acid (42) were isolated from the leaves and showed anti-metabolic and antiviral activites (Table 4) [48].
 Table 4 Anti-metabolic and antiviral activities of compounds (39-42) in Vero cells against CHIKV, and in a DENV RdRp and WNV RdRp assay (µM).
Table 4 Anti-metabolic and antiviral activities of compounds (39-42) in Vero cells against CHIKV, and in a DENV RdRp and WNV RdRp assay (µM).
|
Cell lines |
39 |
40 |
41 |
42 |
|---|---|---|---|---|
|
Vero cells (CC50) cells cecells |
> 420 |
>420 |
2.5 |
19.3 |
|
CHIKV (EC50) |
30 |
>30 |
2.7 |
25 |
|
DENV RdRp (IC50) |
23 |
>23 |
2.2 |
10.5 |
|
WNV RdRp (IC50) |
30 |
>30 |
2.7 |
12.7 |
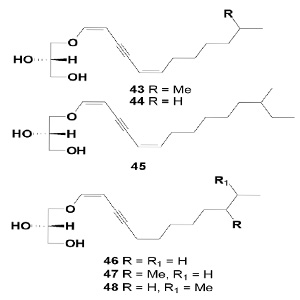
Acetylenic enol ethers of glycerols, including bioactive glycerolipids (43-48), have been isolated from a sponge of the genus Petrosia. Compounds 1 and 3 exhibited weak cytotoxicity against a human leukemia cell-line (K-562) [49]. Bioactivities of glyceryl enol ether compounds (43, 44 and 46), of the yne-diene series, exhibited weak cytotoxicity against the human leukemia cell-line K-562 (LC50 9, 57, 29 µg/mL, for (43, 44 and 45, respectively), while (46-48), possessing the yne-ene group, were less active (LC50 > 100 µg/mL).
Antitumor Sterols and Derivatives:
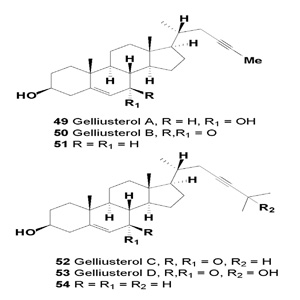
A few species of acetylenic sterols have been discovered, and tested on anticancer activity. Acetylenic sterols, gelliusterol A (26,27-bisnorcholest-5-en-23-yn-3β,7a-diol), its corresponding 7-ketone, gelliusterol B (26,27-bisnorcholest-5-en-23-yn3β-ol-7-one), and gelliusterols C (cholest-5-en-23-yn-3 β-7- one) and D (cholest-5-en-23-yn-3β,25-diol-7-one), were isolated from an unidentified species of sponge, Gellius sp. [50].
Biological evaluation of gelliusterols A (49), B (50), and C (52) was performed on cancer cell lines P-388, HT-29, A-549, DU- 145, and MEL-28. Gelliusterols A and B exhibited moderate activity, with IC50 values greater than 1 µg/mL. An activity of 0.5 µg/mL was observed with gelliusterol C against HT-29, while the other cell lines gave IC50 values above 1 µg/mL. The quantity of gelliusterol D (53) and cholest-5-en-23-yn-3β-ol (68) was insufficient for biological testing [50-52]. Steroids containing an atypical acetylenic unit as a component of the side chain have been obtained from extracts of the sponge Calyx nacaaensis, where 26,27-bisnorcholest-5-en-23-yn-3β-ol (51) and (54) were minor components [51].
A few acetylenic phytosterols were found in plant species. The first acetylenic sterols (55-59) from Gymnostemma pentaphyllum were isolated by Akihisa and co-workers in 1989 [53]. G. pentaphyllum belongs to the Cucurbitaceae family, which is widely distributed in the south of the Qinling Mountains and the Yangtze River in China. G. pentaphyllum is rich in sterols, gypenosides, polysaccharides and flavonoids.
These components have various activities including antitumor, cholesterol-lowering and hypoglycemic activities, anti-aging, anti-inflammatory, and cardiovascular effects [54-57]. Compounds (55 and 56) have similar structures to gelliusterols.
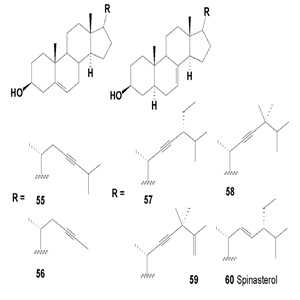
Acetylenic sterols having the same structure (except for the side chains) as the anticancer agent spinasterol (60) were isolated from Pueraria root (Pueraria mirifica from Thailand and Pueraria lobata from Korea) [58]. These plants are used as a rejuvenating folk medicine in Thailand and China. The ethanol extracts had significant antiproliferative effects on breast cancer cell lines, including MCF-7, ZR-75-1, MDA-MB-231, SKBR- 3, and Hs578T. Spinasterol inhibited the growth of some breast cancer cell lines (MCF-7, MDA-MB-231) in a dose- and time dependent manner, as well as the growth of ovarian (2774) and cervical cancer cells (HeLa) [59].
Spinasterol has also been reported in Cucurbita moschata, Conyza blinii, Gypsophila oldhamiana, Gordonia ceylanica, and Acacia cedilloi [59-63]. The anticancer activity of spinasterol was demonstrated in vivo in studies that showed that it greatly decreased the incidence of skin tumors without cocarcinogen or co-tumor promoter activities [64].
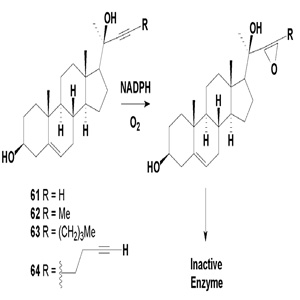
Figure 1:Steroid derivatives with acetylenic side chains as substrates of P-450scc which would generate a reactive species in the active site, thus leading to suicide inhibition of the enzyme.
Several synthesized acetylenic steroids (61-64) having the same structure, except for the side chains, are excellent inhibitors of P-450scc, although they appear to inactivate the enzyme in a manner distinct from the action of acetylenes on the microsomal enzyme [65]. Incubation of (69-72) with P-450scc in the presence of electron donors and oxygen led to a time-dependent absorbance decrease in the Soret region. This absorbance decrease was found to be dependent on the presence of adrenodoxin, adrenodoxin reductase, NADPH, and oxygen. The proposed mechanism of P-450scc inhibition is shown in Figure 1. Additional activities for (61-64) steroids are found as prostaglandin-E2 9-reductase inhibitor.
Cytotoxic Carotenoids:
Three carotenoids with an acetylenic unit, named diadinochrome A (65) and B (66), and diatoxanthin (67) were isolated from the freshwater red tide organism Peridinium bipes (Dinophyceae) [66]. Diadinochrome A was shown to be cytotoxic to HeLa cells, while two other compounds exhibited anti-carcinogenic activity.
Extracts of Peridinium bipes exerted an inhibitory effect on the growth of Microcystis aeruginosa [67-69]. Tsushima and coworkers [68] studied 51 carotenoids, which including carotenoids with acetylenic unit(s): amarouciaxanthin B (sidnyaxanthin), crassostreaxanthin A, diatoxanthin, halocynthiaxanthin, heteroxanthin, mytiloxanthin, mytiloxanthinone, pectenol A and B, and pectenolone. Acetylenic carotenoids showed different cytotoxic activity against Raji cells (human neoplasm). Quantitative carotenoid analysis of the microalga Euglena viridis revealed the presence of β,β-carotene (5% of total carotenoids), mixed with some β,e-carotene, the β,e-carotene derived siphonein (siphonaxanthin 19-dodecenoate, 8%), the allenic neoxanthin (4%), and acetylenic carotenoids > 86% [70-73].
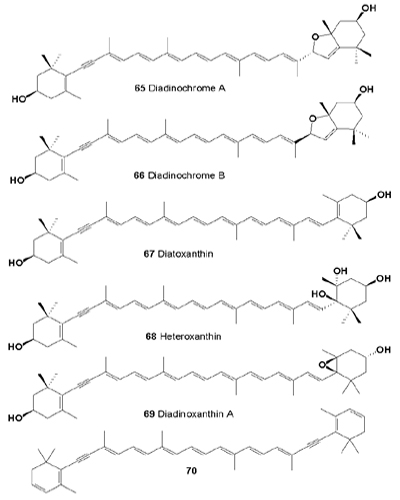
Those included the mono-acetylenic diatoxanthin (67) (major, 61%), diadinoxanthin (69, rearranged to diadinochrome, 12%), heteroxanthin (68, 1%), and the diacetylenic 3,4,7,8,3',4',7',8'-octadehydro-β,β -carotene (70, 6%). The significance of the presence of siphonein and diacetylenic carotenoids for algal chemosystematics was briefly discussed.
Heteroxanthin was also found in Euglena gracilis, and in some species of Xanthophyceae [71-73]. The principal crystallizable xanthophylls of Tribonema aequale were diatoxanthin, heteroxanthin, and diadinoxanthin [74]. Carotenoids of two members of the Raphidophyceae (chloromonads), Gonyostomum semen and Vacuolaria virescens, and of two tentative members of the same class (Chattonella japonica and Fibrocapsa japonica) were analyzed [75].
Group I (G. semen and V. virescens) showed a similar carotenoid pattern, comprised of diadinoxanthin (54-60% of total carotenoids), dinoxanthin (8-17%),β,β-carotene (7%), and heteroxanthin (7%), as well as neoxanthin (G. semen, 3%), an epoxidic monoacetate (G. semen, 12%), an epoxidic carotenol, possibly 9'-cis-diadinoxanthin (V. virescens, 8%), an epoxidic diacetate (V. virescens, 2%) and vaucheriaxanthin 3,19-diacetate (V. virescens, 8%). Characteristic features common to the carotenoids encountered are a high proportion of epoxidic carotenoids (78-86%), allenic carotenoids (24-82%), acetylated carotenols (18-81%), and acetylenic carotenoids (61-67%; Group I only). The xanthophycean cultured alga Pleurochloris meiringensis contains heteroxanthin, diadinoxanthin and β-carotene [76]. Carotenoids extracted from freshwater red tide plankton were shown to include β-carotene (8%), peridinin (26%), dinochrome A (14%), dinochrome B (3%), dinoxanthin (2%), diadinochrome A (65, 3%), diatoxanthin (67, 7%), and 13'-cis-7',8'-dihydro-neoxanthin-20'-al 3'-β-lactoside (4.7%). The isolated carotenoids: β-carotene, dinochrome A, dinochrome B, dinoxanthin, diadinochrome A, diatoxanthin, and 13'-cis-7',8'-dihydroneoxanthin-20'-al 3'-β-lactoside have shown cytotoxic activities against some mouse tumor cell lines [77].
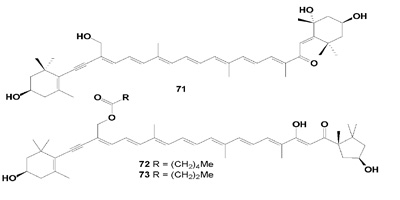
The marine sponge, Prianos osiros from Pohnpei, gave a new cytotoxic acetylenic carotenoid, 3,3',5,19'-tetrahydroxy- 7',8'-didehydro-?,e-carotene-8-one (71), which was cytotoxic toward cultured human colon tumor cells, HCT 116 (IC50 4.4 µg/mL) [78]. Two new carotenoids, the neoplasm inhibitors, 19-hexanoyl-oxymytiloxanthin (72) and 19-butanoyloxy-mytiloxanthin (73), have been isolated from the marine sponge Phakellia stelliderma collected in Okinawa. Both compounds showed mild cytotoxic activity against P388 mouse leukemia cells [79].
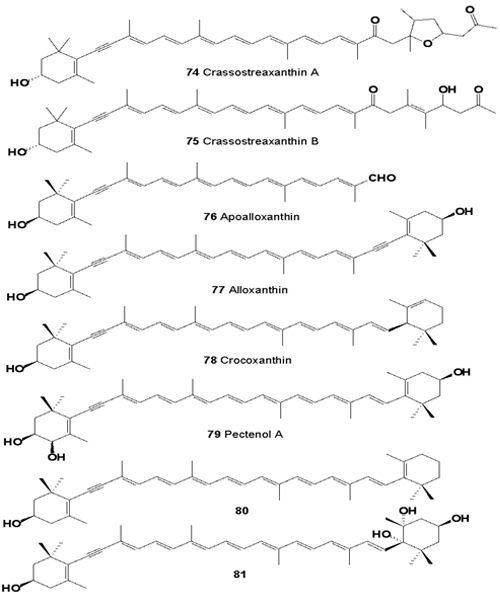
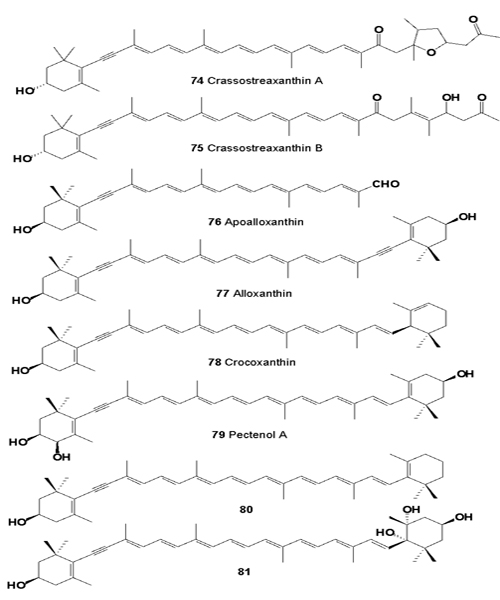
Cytotoxic carotenoids, named crassostreaxanthins A (74), and B (75), and apoalloxanthinal (76), were isolated from the oyster Crassostrea gigas (Ostreidae), and apocarotenoid was isolated (80) from the marine shellfish Mytilus coruscus [80,81].
Acetylenic carotenoids (66, 68, 77, 78, 79, 7,8-didehydro-β- cryptoxanthin 80), and 6-epi-heteroxanthin have been isolated from three species of corbicula clams, Corbicula japonica, C. sandai, and Corbicula sp. (Chinese freshwater corbicula clam) [82].
Total carotenoid content of the muscle of Peronidia venulosa and Corbicula fluminea, and of the gonad of Atrina pinnata and Chlamys farreri ranged from 2.5 to 6.8 mg per cent, values that are relatively higher than those of other shellfishes. The growth of HeLa cells by β-carotene, cynthiaxanthin, astaxanthin and halocynthiaxanthin, NCI-H87 cells by β-carotene, astaxanthin, cynthiaxanthin, and halocynthiaxanthin, HT-29 cells by β-carotene, cynthiaxanthin, mytilo- xanthin and halocynthia- xanthin, and MG-63 cells by β-carotene, cynthiaxanthin, astaxanthin, cantha-xanthin and halocynthiaxanthin were significantly reduced [83].
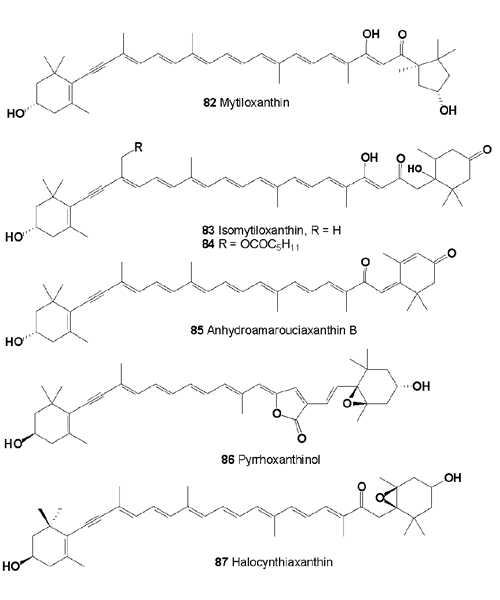
Several cytotoxic acetylenic carotenoids (67, 68, 77, 79), isomytiloxanthin (83), 19'-(hexanoyloxy)-isomytiloxanthin (84), hydroamarouciaxanthin B (85), pyrrhoxanthinol (86), and halocynthiaxanthin (87) have been identified from muscle of Mytilus edulis [84-88]. Diatoxanthin, alloxanthin, and pectinolone from Pectene maximus and Patinopectene yessoensis, pectinols A and B from Mytilus coruscus, crassostreaxanthins A and B from Crassostrea gigas, and a series of carotenoids with a 5,6-dihydro-β-end group from Fushinus perplexus have been reported as the principal carotenoids in marine shellfish [89-92]. Carotenoids in eight species of freshwater and sea mollusks from Russia were investigated [93]. Alloxanthin, mytiloxanthin, isomytiloxanthin, halocynthiaxanthin ether from Modiolus modiolus, and Crenomytilus grayanus; alloxanthin, and mytiloxanthin from Mytilus galloprovincialis; alloxanthin, mytiloxanthin, isomytiloxanthin, halocynthiaxanthin ether, and pectenolon from Mizuhopecten yessoensis have been isolated [93,94].
From the calyx and arms of Lamprometra klunzingeri (family Mariametridae, class Crinoidea, Echinodermata), collected in the Red Sea, the cytotoxic carotenoids diadinochrome (65), alloxanthin (77), cynthiaxanthin, pectenoxanthin, and asterinic acid (88) have been isolated [95].
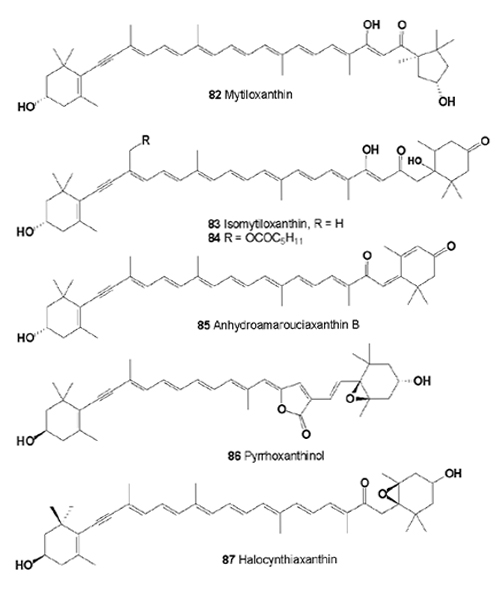
Asterinic acid (88) was found in Echinoderms from the Adriatic Sea: Coscinasterias tenuispina, Marthasterias glacialis, Paracentrotus lividus, and Sphaerechinus granularis [96]. The cytotoxic acetylenic carotenoids diatoxanthin, and alloxanthin were present in the gonads of Australian and Japanese species of the echinoids Heliocidaris erythrogramma and H. tuberculata; in the sea urchin Pseudocentrotus depressus; in Peronella japonica; in seven species of sea-urchins, belonging to the orders Cidaroida, Echinothurioida, Diadematoida, and Arbacioida, as well as pectenolone (89) and 4-keto-cynthiaxanthin (90) [97-100].
Asterinic acid was isolated from Asterias rubens, Acanthaster planci, Coscinastrias acutispina, Leiaster leachii, Asterias amurensis, Ophidiaster ophidianus, Asterina panceri, Asteropecten aurantiacus, and Marthasterias glacialis [101-106]. Mytiloxanthin was found in Ophiocomina nigra; and derivatives of the cytotoxic acetylenic carotenoids, (3S,4S,3'S,5'R)-4-hydroxymytiloxanthin, (3S,4S,3'S,4'S)-4,4'-dihydroxy-diatoxanthin, (3S,4S,3'S,4'S)-4,4'-dihydroxyalloxanthin, (3S,3'S,4'S)-4-keto- 4'-hydroxydiatoxanthin, and (3S,3'S,4'S)-4-keto-4'-hydroxyalloxanthine were isolated from the starfish species Asterina pectinifera and Asterias amurensis [107,108].
The carotenoid content of the seven species of sea cucumber (Stichopus japonicus, Holothuria leucospilota, H. moebi, and H. pervicax of the order Aspidochirotida, Cucumaria japonica, C. echinata, and Pentacta australis of the order Dendrochirotida) was reported [109]. β-Carotene, β-echinenone, canthixanthin, phoenicoxanthin, and astaxanthin were common in all the sea cucumbers examined. Alloxanthin, diatoxanthin, and pectenolone were isolated as minor carotenoids. The bluish violet pigment in the dorsal skin of Asterias rubens was isolated as an amorphous powder [110].
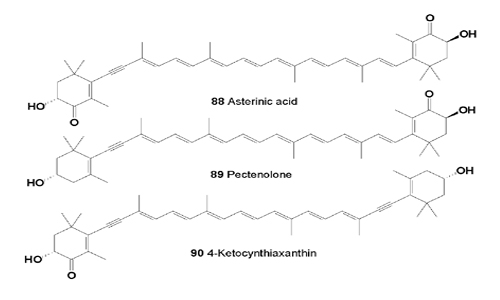
Asterinic acid (100) and four derivatives of alloxanthin, diatoxanthin, and mytiloxanthin were isolated and their structures elucidated. More detailed information on the carotenoids and other bioactive compounds was also reported [111-113].
CONCLUSION:
Intensive searches for new classes of pharmacologically potent agents produced by bacteria, cyanobacteria, micro- and macroalgae, marine and freshwater invertebrates, plants, and fungi have resulted in the discovery of dozens of compounds possessing high cytotoxic activities. However, only a limited number of them have been tested in pre-clinical and clinical trials. One of the reasons is a limited supply of the active ingredients from natural sources. However, the pre-clinical and clinical development of many terrestrial and aquatic derived natural products into pharmaceuticals is often hampered by a limited supply from the natural source.
CONFLICT OF INTEREST:
The author confirms that this article content has no conflict of interest.
REFERENCES:
-
Kuklev DV, Domb AJ and Dembitsky VM. (2013). Bioactive acetylenic metabolites. Phytomedicine. 20, 100-115.
-
Dembitsky VM and Maoka T. (2007). Allenic and cumulenic lipids. Prog. Lipid Res. 46, 328-375
-
Whitehead R. (1999). Natural product chemistry. Ann Rep Prog Chem. 95B, 183-205.
-
Silva FL, Fischer DCH, Tavares JF, Silva MS, et al. (2011). Compilation of secondary metabolites from Bidens pilosa L. Molecules. 16, 1070-1102.
-
Lam JWY and Tang BZ. (2005). Functional Polyacetylenes. Acc Chem Res. 38(9), 745-754.
-
Bohlmann F. (1973). Naturally occurring acetylenes. Phytochemistry. 3, 112-131.
-
Christensen LP. (1998). Biological activities of naturally occurring acetylenes and related compounds from higher plants. Recent Res Develop Phytochem. 2, 227-257.
-
Cadiot P and Chodkiewicz W. (1969). In: Chemistry of Acetylenes. Viehe, H.G. Ed., Marcel Dekker, New York, 597-647.
-
Matsumoto A, Katsuya H, Matsumoto T and Tokuda H. (1991). Antitumor polyacetylene extraction from plants. Japan Kokai Tokyo Koho, Japanese Patent: JP 03287532 A2 19911218 Heisei. 4.
-
Ionkova I and Alferman A. (2000). Use of DNA for detection and isolation of potential anticancer agents from plants. Farmatsiya (Sofia). 47, 10-16.
-
Nakamura N and Nemoto M. (1997). cis-Dehydromatricaria ester concentration in plant and its leaching of Solidago altissima L. Zasso Kenkyu. 41, 359-361.
-
Lu T, Menelaou MA, Vargas D, Fronczek FR, et al. (1993). Polyacetylenes and diterpenes from Solidago canadensis. Phytochemistry. 32(6), 1483-1488.
-
Mell CD. (1937). Interesting sources of natural dyestuffs. Textile Colorist, 59, 482-484.
-
Winarno H, Ohashi K and Shibuya H. (2003). Chemical study on the parasitic plant Scurrula atropurpurea (loranthaceae), an Indonesian medicinal plant. Fukuyama Daigaku Yakugakubu Kenkyu Nenpo. 21, 13-29.
-
Fu X, Abbas SA, Schmitz FJ, Vidavsky I, et al. (1997). New acetylenic metabolites from the marine sponge Pellina triangulate. Tetrahedron. 53, 799-814.
-
Onken D and Heublein D. (1970). Acetylenated steroids. Pharmazie. 25, 3-9.
-
Lee HI, Yoo BS, Yoo MA and Byun SY. (2007). Korean J. Chem. Engineer. 24, 655.
-
Mo Z. (2003). Diss. 172, Hong Kong University of Science and Technology Hong Kong, Peop. Rep. China, Diss. Abstr. Int. 63B, 4453.
-
Wang C, Zhao X, Mao S, Wang Y, et al. (2006). Neurological Res. 28, 436.
-
Razmovski-Naumovski V, Huang THW, Van Hoan Tran G, Li Q, et al. (2005). Phytochem. Rev. 4, 197.
-
Nitsche H. (1973). Heteroxanthin in Euglena gracilis. Archiv Mikrobiol. 90(2), 151-155.
-
Oono M, Kikuchi K, Oonishi S, Nishino H, et al. (1995). Anticancer agents containing carotenoids. Japan Kokai Tokkyo Koho, 5 pp. Japanese Patent: JP 07101872 A2 19950418 Heisei. Application: JP 93-248267 19931004.
-
Nishino H. (1998). Cancer prevention by carotenoids. Mutation Research. 402, 159-163.
-
Matsuno T, Maoka T and Hiraoka K. (1981). A new acetylenic carotenoid from sea mussels. Nippon Suisan Gakkaishi, 47, 143.
-
Campbell SA, Mallams AK, Waight ES, Weedon BCL, et al. (1967). Pectenoxanthin, cynthiaxanthin and a new acetylenic carotenoid, pectinolone. Chem. Commun. 941-942.
-
Tsushima M, Maoka T and Matuno T. (2001). Structure of carotenoids with 5,6-dihydro-a-end groups from the spindle shell Fushinus perplexus. J. Nat. Prod. 64, 1139-1142.
-
Tsushima M, Amemiya S and Matsuno T. (1993). Comparative biochemical studies of carotenoids in sea - urchins. II. The more primitive sea - urchins belonging to the orders Cidaroida, Echinothurioida, Diadematoida and Arabacioida. Comp. Biochem. Physiol. 106B, 729-735.
-
Francis GW, Upadhyay RR and Jensen SL. (1970). Animal carotenoids. 4. Carotenoids of Asterias rubens. Asterinic acid. Acta Chemica Scandinavica, 24, 3050-3052.
-
De Nicola MG. (1959). Composition of new keto carotenoids related to the metabolism of astaxanthin in starfish. Bollett. Sedute Accad. Gioenia Sci. Nat. Catania. 5, 201-214.
-
Goodwin TW. (1969). Pigments in echinodermata. Chem Zool. 3, 135-147.