Information Links
Related Conferences
Previous Issues Volume 6, Issue 1 - 2022
An Update on Mechanistic Modes in AGEs Stimulated & ER and Inflammatory Stress-Modulated Control of the GLUT4 expression (SLC2A4 promoted) and Atherogenesis in Diabetes Mellitus-A Narrative Review
Kulvinder Kochar Kaur1,*, Gautam Nand Allahbadia2, Mandeep Singh3
1Dr. Kulvinder Kaur Centre for Human Reproduction, Punjab, India
2Ex-Rotunda-A Centre for Human Reproduction, Mumbai, India
3Consultant Neurologists, Swami Satyanand Hospital, India
*Corresponding author: Dr. Kulvinder Kochar Kaur, MD, Scientific Director, Dr. Kulvinder Kaur Centre for Human Reproduction 721, GTB. Nagar, Jalandhar-144001, Punjab, India, Tel: 91-181-9501358180, 91-181-4613422, Fax: 91-181-4613422, ORCID: 0000-0003-1473-3419; Email: [email protected]
Received Date: November 17, 2022
Published Date: December 16, 2022
Citation: Kaur KK, et al. (2022). An Update on Mechanistic Modes in AGEs Stimulated & ER and Inflammatory Stress-Modulated Control of the GLUT4 expression (SLC2A4 promoted) and Atherogenesis in Diabetes Mellitus-A Narrative Review. Mathews J Cytol Histol. 6(1):21.
Copyrights: Kaur KK, et al. © (2022).
ABSTRACT
Earlier we had reviewed the role of herbal treatment, epigenetic modes, other mechanistic pathophysiological roles like macrophage polarization, epigenetics, role of adipokines &treatment using role of empagliflozin Extracellular Vesicles (ECV’s), gut microbiota (GM) in Diabetes mellitus (DM) here we concentrated on more work on ER stress, inflammation, AGE’s, Glut4 regulation by its gene. In the last 20 years complicated besides elegant pathways implicated in endoplasmic reticulum (ER) along with inflammatory stress reactions have been illustrated regarding taking part in the generation besides propagation of different diseases inclusive of Diabetes mellitus (DM). These pathways possess different actors taking part pointing to a complex crosstalk amongst ER along with inflammatory stress. Regarding DM, both ER and inflammatory stress are implicated in elimination of glycaemic regulation besides the generation of degenerative complications. Moreover, hyperglycemia enhanced the formation of advanced glycation end-products (AGE), that in turn further triggers the ER along with inflammatory stress, aiding in the inimical glycaemic homeostasis along with escalating the formation of DM complications. Here we conducted a narrative review utilizing search engines PubMed, Google Scholar; Web of Science; Embase; Cochrane review library utilizing the MeSH terms like AGEs; ER stress; inflammation) glucose transporter number 4(GLUT4); soluble carrier family 2 members 4(SLC2A4); glycaemic homeostasis; cardiovascular disease (CVD); atherogenesis from 1990-2022 till date. We found 300 articles out of which we used 130 articles for this review. Here we have highlighted role of AGEs-stimulated ER along with inflammatory stress in the control of GLUT4 expression besides atherogenesis in the form of commonest etiology regarding CVD generation, mortality along with morbidity in DM. Furthermore recently small molecule hampering agents that target these ER stress have been generated from screening products occurring naturally in nature along with synthetic substances, which have undergone clinical trials in animal models along with humans might be the drugs for future treatment of DM besides melatonin, nutraceuticals need further evaluation besides targeting NLRP3 inflammasome.
Keywords: DM, ER Stress, Hyperglycemia, AGE, GLUT4, CVD, Atherogenesis.
INTRODUCTION
Diabetes mellitus (DM) represents a Public Health problem since the numbers of impacted adults are anticipated to reach 230 million by 2045 [1]. DM comprises a metabolic condition with the properties of hyperglycemia that is not appropriate. DM might take place secondary to absence of insulin liberation or type1 DM (T1D) or type2 DM (T2D), basically secondary to insulin resistance (IR) [2]. Nevertheless, while T2D individuals generate dysfunctional insulin liberation, T1D individuals might further generate IR in the form of a complicated outcome obtained subsequent to crosstalk amongst age, sex, pregnancy ethnicity, pregnancy, disease period, escalated life expectancy, genetic proneness, epigenetic alterations, glucotoxicity, lipotoxicity, overweight along with aggravated insulin treatment [2]. Besides glycemic conditions chronic hyperglycemia damages different organ systems resulting in generation of various complications, thus facilitating a 2-4 times enhancement of the risk of cardiovascular disease (CVD) [3].
In case of DM, various metabolic modes noticeably result in activation of intracellular pathways which escalate the formation of advanced glycation end-products (AGEs), hence escalating plasma quantities of AGEs has become an emblem of dysregulated DM [4]. In turn AGEs aid in the IR along with glycemic regulation becoming inimical [5], as well as aid in the generation of chronic complications of DM inclusive of CVD [6]. The receptor for AGEs (AGER-advanced glycation end-products particular receptor-earlier known as RAGE) modulates different biological actions of AGEs, inclusive of the activation of endoplasmic reticulum (ER) stress along with the inflammatory stress pathways [6].
Taking into account that AGEs can take part in the dysfunction of glycemic homeostasis along with the generation/propagation of chronic complications of DM as well as taking into account that these actions implicated, ER stress along with inflammatory modulated pathways. Having reviewed role of herbal treatment, epigenetic modes, along with other mechanistic pathophysiological roles like macrophage polarization, epigenetics, role of adipokines &treatment using role of empagliflozin extracellular vesicles (ECV), gut microbiota(GM) in DM [ 7-21 ], besides in Nonalcoholic fatty liver disease (NAFLD) generation here we review the controlling of these on expression of solute carrier family 2, promoted glucose transporter number 4(GLUT4), in the form of a marker of changed glycemic homeostasis [22], along with on atherogenesis, resulting in a significant induction of the generation/ propagation of (CVD[3].
PATHOGENESIS OF DM AS WELL AS ITS COMPLICATIONS
Insulin resistance (IR)
Insulin resistance (IR) implicates a complicated along with interference in the insulin signaling pathway, ultimately ending in dysfunctional utilization of glucose by muscles along with adipose tissue (ATs), aiding in the dysfunction of the glycemic regulation [22,23]. Moreover, IR exaggerated hyperglycemia by facilitating glucose output from the liver by escalating gluconeogenesis. The glucose overgeneration is unable to neutralize this complementary hyperglycemia, while in a non-resistant manner results in escalated lipogenesis which gets modulated by sterol regulatory element binding protein 1c(SREBP1c), that exaggerates IR as well as facilitates a proatherogenic stage [24]. Further, plasma glucose quantities get determined by crosstalk amongst modes stimulated by IR along with hyperinsulinemia in case of one or greater variable organs.
Uptake of glucose in muscles along with ATs are based on the insulin-responsive glucose transporter GLUT4 that gets encoded by the soluble carrier family 2 member 4 (SLC2A4) gene, that got cloned 3decades back [25]. Subsequently, it got clarified that GLUT4 possesses a foundational part in plasma glucose clearance dysfunction as well as glycemia homeostasis [25]. Moreover, changes in expression of GLUT4 have been correlated with glycemic regulation, with decreased GLUT4 taking part in susceptibility to hyperglycemia. In view of that here SLC2A4/ GLUT4 control of muscles GLUT4 along with ATs is reviewed in the form of a marker of glycemic regulation in DM. Conversely, combinations of IR along with compensatory hyperinsulinemia (in case of T2D) or hyperinsulinemia induction by insulin therapy (in case of T1D) have further been revealed as possessing the capacity of resulting in injury to different organ systems. This causes generation of health complications that result in disability in addition to those that are life-threatening, maximum of which are microvascular (retinopathy, nephropathy, neuropathy) besides macrovascular (CVD, cerebrovascular disease, as well as peripheral vascular disease [3].
IR along with hyperinsulinemia has been correlated with Polycystic ovary syndrome (PCOS), Non-alcoholic fatty liver disease (NAFLD), some kinds of cancer, sleep apnea, along with, in particular, hypertension as well as CVD [26]. Associated with that various conditions correlated with IR have been believed to be the risk factors for hypertension as well as CVD generation inclusive of greater atherogenic lipoprotein profile, escalated plasma plasminogen activator inhibitor-1 (PAI-1), escalated sympathetic nervous system (SNS) action along with endothelial impairment [26]. Additionally, a proinflammatory profile besides activation of endoplasmic reticulum (ER) stress in cells as well as Oxidative stress (OS) is seen in a complicated association resulting in a sinister vicious cycle [27].
GLUT4 expression along with glycemic regulation
The glucose transporter protein GLUT4 is from a family of proteins implicated in glucose facilitative diffusion across the plasma membrane (PM) [28]. GLUT4 represents the lone insulin-sensitive glucose transporter whose expression takes place in skeletal muscle as well as ATs where it is implicated in the insulin-stimulated glucose uptake. In case of myocytes along with adipocytes, insulin binding to its receptor stimulates the excellent intracellular signals sorting which ultimately ends in a storage vesicle GLUT4 translocation to the PM. Subsequent to docking along with fusion processes GLUT4 quantities enhance in the PM, escalating the glucose influx. In view of greater glucose utilization by these cells, the gradient of diffusion persistently facilitates the influx of substrates. Interference with the insulin stimulus results in internalization of GLUT4, which restores the glucose uptake toward the baseline quantities [25]. The GLUT4 modulated escalated glucose uptake in muscles along with ATsis a basic mode implicated in blood glucose clearance in particular in the post-prandial period. Hence various researchers have evaluated the control of the SLC2A4 gene expression that encodes GLUT4 proteins for acquisition of greater insight regarding insulin-modulated plasma glucose clearance [29].
Dysfunction of the insulin signaling transduction possesses the capacity to interfere with GLUT4 translocation to the PM in particular in case of acute conditions where full cellularGLUT4 quantities remain unaltered. Nevertheless, in case of chronic insulin-resistant stages, a reduction of full cellular GLUT4 quantities is presently seen which definitely aids in reduction of GLUT4 at the PM in reaction to insulin. Despite taking into account a conserved transduction system, once the reduction of intracellular quantities of GLUT4 occurs the ultimate quantity of GLUT4 would be decreased [30]. This highlights the considerable importance of the suppression of Slc2a4 gene expression along with subsequent decreases in GLUT4 protein in both IR as well as DM state. Furthermore, the part of Slc2a4/ GLUT4 expression in glycemic regulation got reinstated in studies in transgenic mice. It has been well accepted that knockout of Slc2a4 results in hyperglycemia, while over-expression of Slc2a4 enhances glycemic regulation in DM mice as well [31], controls that were directly correlated with the quantities of GLUT4 at the PM which was independent of alterations in insulin signaling. These outcomes obtained rehighlight the significance of control of SLC2A4 gene expression in glycemic regulation hence making this gene suitable in the form of favourable therapeutic target for generation of pharmacologic treatment for IR along with DM [22].
Currently, various transcription factors have been detailed as implicated in the expression of SLC2A4 /Slc2a4 (human/murine genes respectively) with maximum of them working in the form of enhancers with occasional them as suppressors. Of relevance, certain of these transcription factors have been correlated with inflammatory as well as/or ER stress, which makes these routes significant modulators of GLUT4 expression-detailed later.
Cellular cholesterol homeostasis along with cardiovascular disease (CVD) generation
Cardiovascular disease (CVD) reflects the commonest etiology of ailments along with mortality and morbidity in DM [32]. Despite, the prevalence of CVD exists in both T1D along with T2D it is of greater botheration in case of T2D that accounts for around 91% of all DM patients.IR along with compensatory hyperinsulinemia, obesity, chronic inflammation lipid metabolism along with events controlled along with aberrations that aid in lipid accrual in the arterial wall facilitate the load of clinical complications correlated with CVD. Furthermore, the existence of other risk factors usually correlated with Metabolic Syndrome (MetS) enhances atherogenesis [33].
In T2D quantitative alterations of plasma lipoproteins take place in the form of primary sequelae of this insulin-resistant stage [34]. Hyperinsulinemia escalates hepatic triglycerides (TG) generation by over-activation of SREBP1c. This transcription factor is located in the ER with its transport to the golgi compartment gets regulated by nutritional along with hormonal signaling. This gets followed by the small fragment possessing the leucine zipper liberated subsequent to 2 proteolytic cleavages which are consecutive, gains entry into the nucleus, that results in induction of trans activation of genes implicated in de novo generation of fatty acids (FA). Furthermore escalated lipolysis facilitates the free flow of free fatty acids (FFA) from AT to liver that aids in greater substrate availability for TG generation. These processes aid in the liberation of very low-density lipoprotein (VLDL), that in the blood is metabolized lesser by the lipoprotein lipase that aids in the formation of hypertriglyceridemia. The dysfunction of triglyceride-rich lipoprotein lipolysis results in decreased production of high-density lipoprotein (HDL). Additionally, little along with dense low-density lipoprotein (LDL) formed as per the escalation of plasma TGs quantities, has the properties of greater atherogenic particles which can freely reach the arterial wall which has greater proneness to oxidation. Monocytes obtained macrophages pick up oxidized LDL that results in the first stage of atherosclerotic plaque generation. The escalated formation of oxysterols goes parallel with the intracellular accrual of cholesterol that stimulates inflammation [35].
The balance amongst supply of cholesterol to the arterial wall cells (basically through manipulated LDL) along with eliminated lipid (through HDL) ensures the quantities of lipid accrual which modulates atherogenesis. Reverse cholesterol transportation is the receptive flux of cholesterol which aids in the excretion into bile as well as faeces that features the main guiding force of efflux of cholesterol from the arterial wall section. Lipid free apolipoprotein A1(APOA1) crosstalks with the phospholipids transporting ATPase ATP binding cassette transporter A1(ABCA1), an acknowledged receptor for APOA1 besides growing HDL (pre-beta HDL) that guides additional cholesterol out of cells by receiving energy for the hydrolysis of 2 ATP molecules. Subsequent to the esterification by the phosphatidylcholine-sterol acetyltransferase (namely lecithin cholesterol acetyltransferase) esterified cholesterol assumes its position in the form of integral part of the escalating hydrophobic core of HDL (alias HDL3 followed by HDL2. Bigger HDL crosstalks with the ATP binding cassette transporter G1 (ABGA1), that exports additional cholesterol along with certain oxysterols. Following this HDL attains the capacity of throwing esterified cholesterol directly into the liver by crosstalk with the receptor which forages alias scavenger receptor class B member 1 or indirectly by transfer of esterified cholesterol to LDL, VLDL, or oxysterols by the action of cholesteryl ester transfer protein. In this latter pathway cholesterol elimination modulation occurs by the uptake of APOB possessing lipoproteins by hepatocytes [34].
Advanced Glycation End-products (AGE)
Hyperglycemia results in stimulation of intracellular Oxidative stress (OS) which is believed to be correlated with full DM- associated complications. Escalated cellular glucose facilitates its metabolism via glycolytic pathway that escalates the electron flow via mitochondrial respiration electron transport chain (ETC), that correlates with the formation of Reactive oxygen species (ROS) in particular superoxide anion. The sequelae of enhanced expression as well as action of poly ADP ribose polymerase(PARP), in the form of adaptive mode that confers protection against DNA injury causes modifications of glyceraldehyde 3 phosphate dehydrogenase (GADPH) by poly ADP ribosylation. This dysfunction of glycolysis flow shifts glyceraldehyde 3 phosphate along with dihydroxy acetone phosphate towards the generation of glucosepane methylglyoxal (MGO), an oxaldehyde which is highly reactive besides crosstalks with proteins, phospholipids along with nucleic acids that result in irreversible generation of Advanced Glycation end-products (AGEs) [36]. In general, in case of normoglycemic situations MGO production takes place at highly slow rates with about 0.05-1% of triose phosphates breaking down to MGO. Additionally, its cellular quantity is lesser In view of its action on the glyoxalase system comprising of glyoxalases 1 as well as 2 which transform MGO into S-D lactosylglutathione as well as D lactate respectively [37]. At the time of hyperglycemia further enhancement of AGEs take place in certain cells via the coactivation of the polyol pathway. Transformation of glucose into sorbitol is catalyzed by aldose reductase, that gets transformed to fructose by sorbitol dehydrogenase activity. High reactivity of fructose with protein exists causes pacey accrual of AGEs apart from pacey elimination of nicotinamide adenine dinucleotide phosphate reduced form (NADPH), the polyol pathway causes dysfunction of glutathione regeneration facilitating OS, that enhances the production of AGEs [36].
There is crosstalk amongst the escalated glucose in blood circulation non-enzymatically with the amino-terminal of lysine along with arginine residues in proteins, or with the amino residues of phospholipids that resulted in the generation of unstable Schiff base. As per the sustenance of hyperglycemia, an Amadori product gets formed along with subsequent to inter along with intramolecular reorganization AGEs get generated inclusive of highly heterogeneous substance. Oxoaldehydes, inclusive of MGO, glyoxal,3-deoxy glucosone along with glycoaldehyde represent intermediate products of this event. Dicarbonyl sugars along with other oxoaldehydes possess greater reactivity in contrast to glucose along with besides being escalated in DM are escalated in Chronic Kidney Disease (CKD), inflammation, besides conditions correlated with OS, that resulted in fast production of AGEs. Furthermore, oxoaldehydes enhance in postprandial period facilitating a fast manipulation of circulating proteins. As sequelae the intracellular as well as extracellular protein structure along with working, impacting the extracellular matrix [ECM], cellular crosstalk, receptor modulated cell reactions in addition to DNA structure. Certain AGEs are extreme fluorophores aiding in determination by fluorophores fluorimetry besides might result in induction of covalent crosslinks [38]. Carboxy methyllysine (CML) pentosidine, pyrraline, MGO dimers, glyoxal dimers, MGO obtained glucosepane methylglyoxal hydroimidazolones, as well as glucosepane, reflect the main species of AGEs which have been correlated with long duration DM complications [39]. AGEs, along with other metabolic pathways changed at the time of hyperglycemia inclusive of polyol pathway, protein kinase C activation, besides that of hexosamine pathways comprise the molecular etiology of cellular injury in DM [33].
Crosstalk of AGEs takes place with the receptor AGER, which represents a multiligand receptor which activates NADPH Oxidase, hence inducing ROS formation. This causes nuclear factor κB (NFκB) activation besides transactivation of numerous genes inclusive of AGER that aid in an inimical vicious cycle. Additionally, there is overlap of AGER signaling with the toll-like receptor (TLR) signaling along with possesses the capacity of binding other ligands inclusive of calgranulins, serum amyloid along with high mobility group protein 1 (like amphotericin) enhance inflammatory stress. The placement of AGER on cell surface along with in intracellular organelles aids in the signaling stimulated by various, ligands along with both intracellular as well as extracellularly generated AGEs [40]. Moreover soluble isoforms of AGER with absence of intracellular domain of indigenous receptor, possesses the capacity of binding AGEs without stimulating intracellular signaling hence quantities of soluble AGER in blood might act as a biomarker conferring protection from risk of DM complications [41].
Conversely, the AGE receptor dolichyl-diphospho oligosaccharides protein glycosyltransferase (DDOST or AGER1) hampers AGER signaling by induction of antioxidant genes ameliorated the actions of AGEs. In T1D, a lesser expression of DDOST in peripheral blood mononuclear cells was observed in contrast to healthy controls subsequent to adjusting for age, sex, utilization of statins, angiotensin-converting enzyme (ACE2) hampering agents along with angiotensin receptor blockers [42]. In this way, the balance amongst AGER along with DDOST might be inimical on attempt of evaluation of AGEs tissue injury in DM.
Advanced Glycation causes dysfunction of lipid metabolism aiding in qualitative along with quantitative changes in plasma lipoproteins. AGEs result in modifications of both phospholipids along with apolipoproteins in the lipoproteins structure, thus interference with its recollection by cellular receptors as well as its metabolism by enzymes along with proteins in the plasma along with lymphatic chambers. VLDL along with chylomicron glycation injures the action of enzyme lipoprotein lipase, glycation of APOB escalates its half-life, aiding in the entry of LDL in the arterial wall. Akin to that glycated LDL gets greater oxidation along with immunogenicity, In view of that gets more easily trapped by arterial macrophages [43]. In case of DM HDL modifications by MGO escalated along with robustly interfering with its formation, half-life along with its working, impacting cholesterol flow as well as reverse cholesterol transportation besides atherogenic characteristics of HDL [44].
Endoplasmic Reticulum (ER) stress along with Inflammation in DM
Three decades back ER stress study got started by assessment of the cell responses to heat shock, inclusive of heat shock proteins (HSP), like heat shock 70 kDa proteins (HSP70) [45]. Subsequent to the original detailing of HSP taking part in cellular homeostasis along with immune function, the properties of a perfect ER stress pathway were unraveled with a pleiotropic part advocated inclusive of taking part in the DM formation [46].
Initially, in case of DM, ER stress was correlated with conferring protection to the βcells of the islets of Langerhans in the pancreas. Inspite of the normal generation of endocrine pancreas in eukaryotic translation initiation factor 2-alpha kinase 3((EIF2AK3otherwise PERK) deficient mice, postnatal activation of ER stress is correlated with escalated cell demise leading to a propagative formation of DM [47]. Subsequently, it was illustrated that inositol requiring enzyme1(IRE) deficient mice revealed mild hypoinsulinemia, in particular following a glucose challenge along with inspite of no change in the histological assessment of the pancreatic islets, a decrease in pancreas mass was detailed [48]. Moreover, at the time of generation of DM in Akita mice, overexpression of ER correlated protein BIP (alias endoplasmic reticulum chaperone BIP or GRP78), DDIT3(DNA damage-inducible transcript3 protein or C/EBP-homologous protein (CHOP10)/GADD153 (growth arrest along with enhanced DNA damage-inducible 153) was seen along with targeted breakdown of Ddit3 gene caused postponement of DM initiation, which validated the role of ER stress in β cellular injury [49].
In addition to that ER stress has further been documented to be associated with the pathogenesis of IR along with DM. The generation of high-fat diet (HFD) induced obesity along with T2DM causes reduction of Hspa5 (heat shock protein family A member 5) null mice (knockout for BIP) [50]. Suppression of numerous unfolded protein responses (UPR)-correlated genes further have been seen in β cells exposed to greater quantities of glucose [51]. Actually, numerous studies have illustrated the implications of UPR in the generation along with propagation of metabolic diseases [rev in 52]. Hence currently it is believed that ER stress takes part in the dysfunction of insulin liberation along with activity. Moreover, ER stress has further been correlated with the generation of degenerative complications [53].
In 2000 it got posited that metabolic dysfunction was correlated with escalated inflammatory action, In view of evolutionary causes [54]. It got illustrated that the formation of obesity was correlated with escalated proinflammatory cytokines by the AT, where definition of obesity was a disease with subclinical inflammatory action [55]. It got validated fast regarding the transfer of proinflammatory action to other regions, hence taking part in the pathogenesis of T2D [53, 54].
Once the molecular modes stimulated by inflammation were getting worked out apparent clarification revealed that numerous of them had shared with the UPR pathway, documenting crosstalk amongst these 2 events. Like the activation of certain UPR constituents like BIP, IRE, TRAF2 (TNF receptor-associated factor2) results in IKKB (inhibitor of nuclear factor κB kinase subunits beta/ alpha) phosphorylation stimulating the activation of NFκB, a significant modulator of the inflammatory action [56]. It is clear now that UPR reflects a proteostatic pathway possesses the capacity of modulating inflammation along with immunity [57].
GLUT4 EXPRESSION ALONG WITH ER STRESS/ INFLAMMATION
Subsequent to it getting pointed that the ER stress activation possesses a significant part regarding the pathophysiology of T2D, evaluation of ER stress regarding the controlling of GLUT4 expression in adipocytes was initiated. Following that it was revealed that ER stress reaction resulted in reduction of Slc2a4 expression at the gene transcription level as well as escalated expression of Ddit3 gene. It has been acknowledged that the DDIT3 protein is a hamperer of the activity along with the expression of the Cebpa gene (CCAAAT enhancer binding protein alpha), with the CEBPA transcription factor being a robust activator of Slc2a4 / GLUT4 expression [58].
Muscle studies were initiated in insulin-resistant mouse C2C12myotubes where a mixture of black soybean peptides enhanced the glucose transport along with GLUT4 translocation with simultaneously hampering the ER stress reaction [59]. Moreover, in rat myotubes, IR induction by glucosamine therapy was documented to cause activation of certain ER stress markers like BIP along with XBP1(X-box binding protein 1) besides escalating activating transcription factor6 (Atf6) gene [60]. Over generation of ATF6 hampers the expression of certain significant enhancers of Slc2a4 expression like myocyte enhancer factor 2 A(MEF2A), PRGC1A (Peroxisome Proliferator-Activated Receptor coactivator 1-alpha or PGC-1α) which reasons out the suppression of Slc2a4/GLUT4. Lastly, it was validated that silencing Atf6 (with utilization of small interfering RNA) was enough for complete avoidance of glucosamine-stimulated actions. These outcomes exquisitely illustrated that ER stress results in IR in case of myotubes through dysfunction of muscles GLUT4 expression by utility of ATF6-based pathway [60]. Subsequent to the initial pointer that inflammation possesses the capacity of hampering GLUT4 expression [53], the group of MachadoUF initiated concentrating on illustrating that control. Initially, it was pointed that (in case of soleus muscle regarding in vivo metabolic situations (like fasting) along with in vitro therapy (like muscle contraction along with incubation with insulin) following Slc2a4 mRNA expression in contrast to Nfkb mRNA expression [61]. Electrophoretic mobility shift assay (EMSA) further illustrated that the NFκB binding activity towards an NFκB binding region in agreement alters in conjunctions with Nfkb mRNA expression [61]. These outcomes robustly pointed out that the inflammatory stimulated suppression of Slc2a4 gene expression implicates an NFκB- modulated transcription action.
NFκB-modulated activity taking part in Slc2a4 mRNA expression got validated further by i) anti-inflammatory action of atorvastatin in case of AT of T2D mice [62], ii) the insulin-sensitizing action of Cannabinoid receptor1(CBR1) in 3T3L1 adipocytes [63], iii) the dosage-based insulin-stimulated escalation of Slc2a4 expression in soleus muscle [64] along with iv) oleic acid as well as linoleic acid fatty acids (FAs) stimulated suppression of Slc2a4 expression in L6 muscle cells [65]. These studies outcomes illustrated that there was an opposite control amongst Slc2a4 gene expression along with NFκB-expression or activity which revealed that the transcription factor NFκB binding possesses a suppressor action on the Slc2a4 gene; which is highly unlikely taking into account that presently NFκB is acknowledged to escalate numerous genes.
Of significance the promoter area of the Slc2a4 gene does not reveal the kappa B binding region in agreement; hence the group of Machado UF evaluated a homozygous sequence residing in-134/-113 areas of the mouse Slc2a4 gene. They validated that both p50, as well as p65 subunits of NFκB, possess the capacity of binding to this area in vivo and in vitro (EMSA) along with in vivo (chromatin immunoprecipitation assay). Furthermore, a reporter gene assay validated that this area is implicated in hampering the transcription of Slc2a4 [62]. Actually, in maximum of their studies regarding inflammatory stimulated control of Slc2a4 expression they have validated the binding action of both p50 as well as p65 subunits of NFκB into this -134/-113 Slc2a4 promoter area [62-65].
Lastly, they illustrated the implication of inflammation along with ER stress in the suppression of Slc2a4/GLUT4 expression in the muscle cells having exposure to palmitate [66]. Acute therapy escalated the BIP, eukaryotic translation initiation factor 2A (EIF2A) EIF2AK3, IRE1 along with TRAF2 protein quantities, along with EIF2AK3 phosphorylation, however, did not stimulate EIF2A along with IKK phosphorylation or escalated XBP1 nuclear quantity. In addition to that acute along with chronic therapy escalated the nuclear quantity of NFκB p65 subunits as well as nuclear binding activity in the Slc2a4 gene promoter. These total outcomes obtained studies illustrated that the palmitate therapy stimulated the activation of UPR like the generation of IRE1-TRAF2-IKK complex that intersects with an NFκB modulated Slc2a4/ GLUT4 along with illustrated an association amongst ER stress besides inflammation in IR [66].
Endoplasmic reticulum (ER), as well as inflammatory stress (with decreased GLUT4 expression), have been seen in insulin-resistant skeletal muscles from women with Gestational Diabetes mellitus (GDM) along with suppression of ER stress by tauroursodeoxycholic acid (TUDCA) or si RNA knockout for IRE1A or BIP along with suppression of muscles GLUT4 resulted in a significant downregulation of activation of ER as well as inflammatory stress, escalated GLUT4 expression as well as glucose uptake [67]. Intriguingly, it was illustrated that continuous preoperative fasting in rats stimulated postoperative ER stress activated by IRE1A as well as suppression of muscles GLUT4 resulting in IR along with hyperglycemia [68]. Lastly, a recent study conducted in hippocampus of DM rats correlated with ER stress activation, BIP, DDIT3, ATF4) along with inflammatory stress (Tumor necrosis factor alpha (TNFα) as well as interleukin-6(IL-6) with decreased GLUT4 expression. Moreover, metformin/donezepil treatment was illustrated in reverting these changes; thus, an attractive approach regarding management of -DM correlated dementia [69].
Atherogenic/Cardiovascular disease (CVD) along with ER stress/Inflammation
Hyperglycemia is acknowledged to be an inimical factor for the production of CVD, DM, obesity along with other metabolic diseases, changed lipid metabolism acts in the form of a significant mode of etiopathogenesis. The modulation of lipid metabolism works in the form of a biomarker along with pathological factor regarding CVD, DM, obesity along with other metabolic diseases, in the case of these situations, dysfunctional cellular homeostasis interferes with appropriate working of the ER, hence ER stress, as well as inflammation, possesses an elemental part regarding the pathogenicity of these particular diseases [70].
A direct association of ER stress exists amongst scavenger receptor A, the platelet glycoprotein 4(FAT, or SCARB3) plasma pathological oxidized low-density lipoprotein with) along with their hampering acts as targeting for avoidance of generation of atherosclerosis [71]. Inflammatory as well as ER stress markers guided by the accrual of free cholesterol- along with 7 ketocholesterol are escalated in atherosclerotic lesions areas having susceptibility for rupturing [35,72].
The chronic ER stress activation is intricately correlated with endothelial impairment as well as atherosclerosis [73]. The free cholesterol accrual in cells, inflammation along with other correlated CV risk factors (hyperhomocysteinemia, saturated fatty acids, manipulated lipoproteins as well as DM) stimulate or are inimical for ER stress [74]. Cholesterol esterification by the sterol O’acetyl transferase (or ACAT) aiding in the storage of esterified cholesterol in a germane inert kind in the cytosol besides pacey bioavailability of free cholesterol. Additionally, avoidance of non-particular free cholesterol flux amongst cell organelles which is in particular toxic towards ER. The assessment of the transcriptome of aortic tissues in case of high-fat diet (HFD) fed Apoe knockout mice illustrated the escalation of 3 main UPR pathways along with overlap of 50 genes implicated in UPR signaling, its adaptability along with apoptosis [75]. Moreover, in case of DM, Apoe deficient mice, in receipt of glucosamine supplementation, illustrated bigger, besides atherosclerotic lesions having greater advancements at the aortic root that illustrated greater immunoreactivity for BIP as well as ENPL (endoplasmin, otherwise GPR94) at the site of lesion tissue was seen [76].
Escalated cholesterol, as well as cholesterol crystals, further initiates the activation of the NLRP3 inflammasome that along with the ER stress activation are in close association which underlies the formation of atherosclerosis [77]. Intriguingly of the modulators of UPR, ER placed ATF6 is akinto SREBP1c, that gets activated by 2 proteolytic cleavages in the golgi chamber [78]. ATF6, despite hampering the activation of the SREBP1c, by itself possesses the capacity of induction of lipogenesis, generating an association amongst UPR along with dyslipidemia has been illustrated [79].
Liver X receptor (LXR) activation by escalating the expression of ABCA1 as well as ABCG1 enhances the cholesterol efflux, ameliorates ER stress along with decreased apoptosis as well as impaired apoptosis efferocytosis in case of atherosclerotic plaques [80]. Pharmacologic hampering of cholesterol esterification along with what stimulates enhanced cholesterol efflux subsequent to treatment with APOA1 mimetics or APOA1 overexpression, ameliorate ER along with inflammatory stress [81]. Other than hampering oxidation along with inflammation, besides stimulating escalated cholesterol efflux, HDL aids in ER stress avoidance [81]. Vascular smooth muscle cells treated with greater quantities of glucose medium stimulates the expression of CD36, cytokines besides markers of calcification along with ER stress, a situation which gets exacerbated on addition of oxidized LDL (oxLDL). These outcomes were validated by observation in carotid plaques derived from DM individuals in contrast to non DM individuals [82].
AGE stimulated along with ER stress/Inflammation-Modulated actions in DM
AGE stimulated Inflammation, OS along with sterol accrual stimulates ER stress with the properties of escalated expression of BIP ENPL, EIF2A as well as ATF6 (cyclic AMP-dependent transcription factor - ATF6 alpha). Moreover, glycation by itself aids in protein misfolding, stimulating UPR in the form of an intracellular protein nuclear quality regulation system [83]. Glycation alters the electrostatic crosstalk along with the hydrophobicity of polypeptide residues which stimulate ubiquitination along with breakdown [84]. Actually, chronically co-existence of AGEs along with ER stress takes place at the time of numerous biological situations indicating their inherent actions in the propagation along with other carbonyl stress correlated complications. Additionally, the complications progression of this close mode gets corroborated by the observation regarding the expression of AGER along with its ligand-calgranulin-S100B enhances its neurological impairment marked by escalated ER stress activation [85].
Endothelial cells proteome which had incubation with greater glucose quantities medium isolated the enrichment of 331proteins having differential expression in contrast to normoglycemic medium. Besides the proteins implicated in glycolysis as well as gluconeogenesis, HSPs correlated with UPR along with proteins refolding, the ubiquitin E2 ligases implicated in the protein breakdown by the proteasomal along with lysosomal system escalated on toxic glycoOS modulated by MGO production [86]. The plasma quantities of tribble homolog3protein (TRB3) [ which gets escalated by hyperglycemia along with ER stress were observed in individuals with T2D along with AGEs, BIP along with TNF, moreover it has a positive association with fasting plasma glucose along with AGEs [87].
AGEs along with GLUT4 expression Control
AGEs have been correlated with the generation along with propagation of DM stimulated degenerative complications in details; nevertheless, we possess minimum knowledge regarding their taking part in glucose homeostasis as canonically they make their appearance only in conditions of hyperglycemia. Nevertheless recently it has been noticed that generation of AGEs can be found in situations of carbonyl stress besides might be derived from external sources in view of which AGEs are considered to be having a crucial part regarding insulin sensitivity/GLUT4/glucose homeostasis that makes a vicious cycle close. Actually, plenty of proof exists about the part of external AGEs, basically from dietary sources in IR along with DM induction in case of animal models along with humans [88].
Regarding humans trials that implicated greater AGEs possessing diet is not feasible in view of ethical reasons; however, studies conducted with human utilization of lesser AGEs possessing diet illustrated attenuation of IR that is partially reasoned out towards the escalated expression of DDOST along with SIRTUIN, in view of their antioxidant, along with anti-inflammatory effects [89].
In vitro studies have illustrated that the direct actions of AGEs regarding insulin sensitivity aided and tissues insulin sensitivity in particular in GLUT4 control along with inflammatory activity. Overexpression of AGER in 3T3L1preadipocytes aggravated adipocyte hypertrophy, while its hampering with utilization of small interfering RNA caused reduction of adipocyte hypertrophy. Moreover, AGER-stimulated adipocyte hypertrophy was correlated with decreased insulin signaling along with glucose uptake, downregulation of GLUT4 along with upregulation of toll-like receptor 2(TLR2) [90]. In addition to that in cultured human preadipocytes adipogenesis was correlated with escalated quantities of CML along with AGER, besides CML was observed to stimulate an AGE- based dyscontrolled inflammatory adipokines [91]. Moreover, a recent study validated that 3T3L1 adipocytes cultured in a pro glycating medium (with MGO or MGO modulated bovine serum albumin) escalated Ager along with decreased Slc2a4 gene expression; apart from bovine serum albumin-MGO decreases glucose uptake [92].
Intriguingly epicardial adipose tissue (EAT), a visceral fat that surrounds the myocardium, that has the probability of being implicated in the initiation/ propagation of coronary artery disease (CAD) has been evaluated in individuals going via open heart surgery. Enhanced EAT was seen to be associated with escalated EAT thickness, decreased GLUT4 expression, adiponectin along with lactoyl glutathione lyase (or glyoxalase1) along with escalated expression of the high mobility group protein B1, TLR, in addition to myeloid differentiation primary response protein (MyD88). These outcomes pointed that in individuals with CAD, AGER might be implicated in facilitating EAT adiposity along with metabolic impairment which is correlated with reduced GLUT4 expression [93].
The initial pointer of primary in vivo AGE action regarding glycemic homeostasis along with GLUT4 expression was revealed in experimental healthy Sprague Dawley rats that had chronic methylglyoxal infusion with use of mini pump. Escalated plasma glucose in this model caused decreased GLUT4 expression along with glucose uptake (AT) along with robust beta cell dysfunction which mimicked a T2D profile [94]. Moreover in healthy Wistar rats chronic delivery of AGE-albumin (x3mths) escalated adiposity along with body weight, besides induction of whole body IR correlated with enhanced inflammatory markers along with, reduction of Slc2a4/ GLUT4 expression in periepididymal adipose tissue [95].
Lastly, the taking part of ER stress along with inflammation in the in vivo along with In vitro AGE action in the Slc2a4/ GLUT4 expression in skeletal soleus muscles of healthy rats [5]. Regarding in vivo assessment, treatment of rats with AGE- albumin, which generated whole body IR along with, reduction of Slc2a4/ GLUT4 expression, escalated nuclear quantities of NFκB (p50) along with escalated cellular quantities of BIP [4]. regarding in vitro assessment, incubation (for2.5-7.5h) of muscles from healthy rats with AGE-albumin was conducted, besides illustrated decreased quantities of Slc2a4/ GLUT4, escalated quantities of BIP, ENPL, along with escalated phosphorylation of IKKA besides IKKB along with nuclear quantities of NFκB (p50 as well as 65). Moreover, EMSA assessment illustrated escalated nuclear proteins NFκB binding in NFκB binding areas of the Slc2a4 promoter [5].
These outcomes illustrated that greater AGE quantities cause dysfunctional glucose homeostasis via a mode implicating ER stress/inflammation modulated suppression of Slc2a4/ GLUT4 expression. Figure 1 illustrates the major modes by which AGE stimulated along with ER stress modulated suppression of GLUT4 expression [96].

Figure 1. Courtesy ref no-95-Advanced glycation end products (AGE)-induced regulation of glycemic homeostasis and glycemic homeostasis: participation of inflammatory and endoplasmic reticulum stress. In adipose and muscle tissues, AGE/CML binding to AGER and TLR activate the ER-stress response, including increased expression and/or activation of BIP, ENPL, XBP1, DDIT3, IRE1 and TRAF2. XBP1 and BIP activate ATF6, which is a repressor of the transcription factor PRGC1A and MEF2A (enhancers of Slc2a4 gene), thus decreasing Slc2a4 expression. On the other, IRE1/TRAF2 activation overlap the inflammatory pathway phosphorylating IKKA/B, and promoting NFKB dissociation and migration into the nucleus, where it plays a potent repressor effect upon Slc2a4 gene expression. Besides, intracellular MGO can contribute to the increase in the activity of proinflammatory cytokines, reinforcing this pathway. These effects on the Slc2a4 gene expression decreases GLUT4 synthesis and plasma membrane content, eventually decreasing insulin-induced glucose uptake, and establishing IR. In turn, IR leads to hyperglycemia, which can cause T2D or worsen pre-existing DM. Symbols are: AGE, advanced glycated end product; AGER, advanced glycation end product-specific receptor, alias RAGE; ATF6, cyclic AMP-dependent transcription factor ATF; BIP, endoplasmic reticulum chaperone BIP, alias GRP78; CEBPA, CCAAT enhancer binding protein alpha; CML, carboxymethyl-lysine; DM, diabetes mellitus; DDIT3, DNA damage-inducible transcript 3 protein, alias CHOP10/GADD153; DM, diabetes mellitus; ENPL, endoplasmin, alias GRP94; ER, endoplasmic reticulum; GLUT4, solute carrier family 2, facilitated glucose transporter member 4; IKKA/B, inhibitor of nuclear factor kappa-B kinase subunits alpha/beta; IR, insulin resistance; IRE1, inositol requiring enzyme-1; MGO, methylglyoxal; MEF2A, myocyte enhancer factor 2A; NFKB, nuclear factor NF-kappa-B; PRGC1A, peroxisome proliferator-activated receptor gamma coactivator 1-alpha, alias PGC1A; Slc2a4, solute carrier family 2 member 4 gene; T2D, type 2 diabetes mellitus; TLR, toll-like receptors; TRAF2, TNF receptor-associated factor 2; XBP1, X-box-binding protein 1. Names and symbols of proteins are in accordance with the UNIPROT database (https://www.uniprot.org/, accessed on 2 November 2021). Parts of the figure were drawn from Servier Medical Art (https://smart.servier.com/, accessed on 2 November 2021).
AGEs along with Atherogenesis/CVD generation
Inflammation possesses a significant in the generation of CVD, that can be initially seen by its part regarding macrophage homeostasis. The maximum enrichment of protein in circulation-namely of albumin possesses escalated proneness towards glycation, besides AGE-albumin escalates the formation of ROS in macrophages, hence primes these cells towards the lipopolysaccharides (LPS) stimulated liberation of proinflammatory cytokines [97] along with stimulating the accrual of oxysterols [98].
In case of non-Diabetic dyslipidemic mice (Apoe knockout mice), AGE-albumin treatment enhanced lipid infiltration in the bovine aortic arch, escalated the expression of Ager, Tnf, Nox4 (NADPH Oxidase) genes besides enhanced the marker of lipid peroxidation (4 hydroxynonenal) along with CML in contrast to animals in receipt of control albumin. Intriguingly treatment with losartan (angiotensin 2 receptor antagonist) ameliorated lipid accrual in addition to the expression of AGE-AGER axis in the aortic arch of animals in receipt of AGE-albumin [99].
The properties of ER stress taking part in the production of CVD have been initially studied in macrophage homeostasis. OS stimulated a decrease in the ABCA protein quantity along with cholesterol efflux to APO1; avoidance of both processes with utilization of aminoguanidine, a substance possessing the antioxidant along with antiglycation properties which decrease ROS generation [100]. Apart from the utilization of a chemical chaperone (namely 4-phenyl butyric acid), that ameliorated protein misfolding caused ABCA quantity reversal [101]. The ER-associated degradation (ERAD) pathway implicated in the breakdown of ABCA1 in addition to the endosomal sorting complex required for transport (ESCRT) besides proteolysis modulated by surface calpains [102].
Macrophages which were in receipt of AGE- albumin (either generated in vitro or derived from subjects having poor DM control) revealed rapid breakdown of ABCA1 in view of the ubiquitin-proteasomal along with lysosomal that are independent of surface calpains [103]. These actions on interference with APO1 modulated cholesterol elimination along with stimulating inflammation continued despite cell rest for long once AGE-albumin was lacking. Additionally, the dysfunctional cholesterol efflux restoration subsequent to the enhanced glycemic regulation pointed towards the reduction of AGE quantities [104]. With the lack of AGER in macrophages with acquisition by AGER silencing or utilization of peritoneal macrophages from Ager knockout mice, the inimical actions of AGE-albumin generated in vitro or derived from subjects having poor DM control were ameliorated. Furthermore, the changed expression of genes implicated in lipid efflux restoration occurred in lack of AGE /AGER signaling [105]. Taking into account the capacity of numerous receptors to crosstalk with AGEs, future studies are essential for the acquisition of greater insight regarding the blockade of AGE /AGER signaling for enhancement of cholesterol balance in the arterial wall.
Diabetic Kidney Disease (DKD), represents a frequent complication of DM which enhances cardiovascular mortality. Even in Chronic Kidney Disease (CKD) prevalence of AGEs are high in view of the Maillard reaction intermediates. Apart from that, carbamoylation takes place in view of dissociation of urea into thiocyanates, whose reaction takes place with proteins. In case of an animal model of uremia carbamoylated albumin interfered with cholesterol efflux that was correlated with ER stress [106]. Albumin carbamoylation in humans with DKD escalates as per the decreased estimated glomerular filtration rate (GFR) along with decreases HDL-modulated cholesterol efflux from macrophages [107]. As per the probable modes implicated it has been documented that AGE-albumin escalated the expression of BIP along with stimulated apoptosis-based on the intracellular calcium escalation in kidney podocytes. Moreover, TUDCA that leads to dysfunction of ER stress by working in the form of a chaperone avoids the cell demise [108]. Additionally, MGO-obtained AGEs were validated to stimulate ER stress (enhanced BIP, ATF4, DDIT3) in case of kidney cells correlated with Inflammation, apoptosis along with mitochondrial function imbalance [109].
EIF2AK3/PERK activation was seen in an animal model to be implicated in AGE-stimulated coronary impairment [110]. Moreover AGE- stimulated ER stress activation was further correlated with exacerbated post-myocardial infarction ventricular arrythmias in Diabetes [111]. Glycated LDL stimulated ER stress in case of bovine aortic endothelial cells, which causes activation of BIP, EIF2AK3 ATF6 that was associated with dysfunctional endothelial-based vasodilation [112]. Furthermore, in human vascular endothelial cells the decreased activity of antioxidant enzyme paroxonase 2 induced by AGEs is correlated with escalated Oxidative, Inflammatory, along with ER stress [113]. Ultimately, an advantageous action of decreasing AGEs quantities for CVD in humans was pointed in the recent CORDIOPREV, in which the decreased circulating AGEs attained by Mediterranean diets determined by the MGO quantities was correlated with higher potential of 4remitting of T2D in individuals with CVD [114]. Figure 2 depicts the major modes in the AGEs stimulated along with ER stress-modulated atherogenesis along with CVD production.

Figure 2. Courtesy ref no-95-Advanced glycation end products-induced atherogenesis and CVD development: participation of inflammatory and endoplasmic reticulum stress. Glycated LDL reaches the arterial intima inducing circulating monocyte chemotaxis. Differentiated macrophages take up modified LDL by the scavenger receptors which induce intracellular cholesterol accumulation. In addition, advanced glycated albumin (AGE-albumin), modified in blood circulation or in the arterial intima is taken up by AGER. This induces the generation of ROS by NOX4 and mitochondrial respiratory chain activation. Moreover, AGEs induce inflammation and ER stress, with increased expression of BIP, ENPL, ATF6, EIF2AK3 and DDIT3 that relates to ABCA1 degradation by the ubiquitin-proteasomal and lysosomal systems. Ultimately, a dramatic decrease in ABCA1 protein content in macrophage foam cells is observed, leading to the reduction in the APOA1 and pre-beta HDL-mediated cholesterol efflux. Although AGEs do not alter ABCA1 mRNA, they reduce the gene expression of ABCG1, by an LXR-dependent mechanism, then compromising the HDL-mediated cholesterol efflux. The accumulation of cholesterol and oxysterols perpetuates the generation of ROS, inflammatory cytokines and AGEs, which culminates in a necrotic core formation, with apoptosis, pyroptosis, plaque instability and rupture. The deleterious effects of AGEs on macrophage homeostasis damage the cholesterol flux to the liver by the reverse cholesterol transport contributing to the independent relation of AGEs with CVD. Symbols are: ABCA1, phospholipid-transporting ATPase ABCA1; ABCG1, ATP binding cassette transporter G-1 AGE, advanced end product; AGER, advanced glycation end product-specific receptor, alias RAGE; APOA1, apolipoprotein A-1; ATF6, cyclic AMP-dependent transcription factor ATF-6; BIP, endoplasmic reticulum chaperone BIP, alias GRP78; CVD, cardiovascular disease; DDIT3, DNA damage-inducible transcript 3 protein, alias CHOP10/GADD153; EIF2AK3, eukaryotic translation initiation factor 2-alpha kinase 3, alias PERK; ENPL, endoplasmin, alias GRP94; HDL, high-density lipoprotein; LDL, low-density lipoprotein; LXR, liver X receptor; NOX4, NADPH oxidase 4; ROS, reactive oxygen species. Names and symbols of proteins are in accordance with the UNIPROT database (https://www.uniprot.org/, accessed on 2 November 2021). Parts of the figure were drawn from Servier Medical Art (https://smart.servier.com/.
AGEs, ER stress along with inflammation in Non-alcoholic fatty liver disease (NAFLD)
With the advancements of ER stress along with inflammation their implications in chronic diseases like DM, CVD, neurodegenerative conditions, Non-alcoholic fatty liver disease (NAFLD), inflammatory bowel disease (IBD), cancers besides others are getting clarified [115]. Remarkable interest has been generated regarding NAFLD in the form of a complication of DM in view of enhanced incidence along with robustness, with ER along with inflammatory stress have been illustrated to be implicated in this condition.
Diabetes mellitus Apoe knockout mice, in receipt of glucosamine supplementation generated intracellular lipid accrual pointing to the formation of NAFLD which was correlated with enhanced quantities of UPR markers BIP, ENPL, DDIT3 along with protein disulfide-isomerase in case of liver tissues [79]. Conversely, the group of Machado UF, documented that T2D mice generated histological signals of Non-alcoholic steatohepatitis (NASH) as per NASH-CRN Pathology Committee System correlated with dysfunctional glucose metabolism in the liver, along with enhanced expression of inflammatory markers TNF, IL-6, as well as NFκB along with escalated binding activity of nuclear proteins NFκB binding in NFκB binding target genes, as determined by the Slc2a2 (soluble carrier family 2 members 2) gene [116].
The P2Y purinergic receptor 12, an adenosine diphosphate responsive G-protein coupled receptor that gets expressed on the surface of platelets (needed for normal platelet collection) gets further expressed by the macrophages in case of liver from mice with cirrhosis as well as Hepatocellular carcinoma (HCC). Ticagrelor stimulated hampering of P2Y purinergic receptor12 escalates tumor cell phagocytosis by macrophages as well as causes induction of anti-tumor phenotype that was correlated with enhanced expression of players of ER stress pathways pointing to activation of UPR [124]. Apart from that, hampering of UPR with TUDCA the prophagocytic actions of ticagrelor, which validated the implications of P2Y purinergic receptor12 modulated macrophage function via the activation of ER stress [117]. Apparently, this is germane regarding the pathogenesis of chronic liver disease as well as cancer.
Enhanced intake of sugars, in particular fructose along with abundant AGE-rich diet are associated with the formation of intracellular AGEs which cause induction of lipogenesis in addition to inflammation in crosstalk with ER stress [118]. Apart from the extravasation of AGEs as well as their crosstalk with RAGE has a correlation with formation besides evolution of different chronic diseases inclusive of steatosis along with CVD [119]. A correlation amongst escalated quantities of different kinds of AGEs along with NAFLD was illustrated in a recent meta-analysis [120]. The estimation of fluorescent AGEs has been regarded to be a probable biomarker for stratifying NAFLD [121].
In to current crosstalk amongst AGEs, ER stress along with inflammation in the production as well as/or propagation of NAFLD in DM gets intriguingly more obvious [122].
POTENTIALTHERAPIES
Role of NLRP3 inflammasome
Role of H2S
Recently we had reviewed the role of nucleotide-binding domain, leucine-rich-repeat containing family, pyrin domain- containing 3 (NLRP3) inflammasome, that is a recent innovation regarding cytoplasmic multiprotein complex implicated in inflammation [123]. The NLRP3 inflammasome possesses NLRP3 along with apoptosis- correlated speck like protein possessing a caspase enrollment domain (ASC) via its N terminal PYD, as well as precursor caspase 1. NLRP3 inflammasome is implicated in numerous diseases inclusive of DM. H2S represents an inimical gas possessing rotten egg aroma. Recently its isolated in the form of a 3rd gas signal subsequent to nitric oxide (NO), as well as carbon monoxide. It possesses numerous biological functions possesses considerably significant part in innumerable diseases inclusive of DM. Recently it has been illustrated that H2S control of the inflammasome aids in various diseases. Hence Zhao, et al. [124], detailed its modes in control of NLRP3 inflammasome in DM for a theoretical manner for further evaluation (Figures 3& 4) [124].

Figure 3. Courtesy [124] Schematic diagram of the NLRP3 inflammasome activation process.

Figure 4. Courtesy [124] Summary of the production of endogenous H2S. CBS: cystathionine-beta-synthase; CSE: cystathionine-gamma-lyase; 3-MST: 3-mercaptopyruvate thiotransferase; 3-MP: 3-mercaptopyruvate; CAT: cysteine aminotransferase.
Role in Atherosclerosis
Atherosclerosis representing a commonest complication of DM, implicated numerous pathological factors. These are inclusive of endothelial impairment, inflammation along with Oxidative stress (OS), which are the corner store of DM as well as atherosclerosis. Despite capacity of DM to facilitate atherosclerosis has been illustrated, a greater insight regarding the underlying mode is key for Identification of innovative targets. NLRP3 inflammasome possesses considerable significant part in DM as well as atherosclerosis. Whereas the variety of its activation modes remains part of its complicated actions regarding the propagation of DM as well as atherosclerosis it further aids in acquisition of deeper insight regarding the targeted modulation of metabolic diseases (Figure 5&6) [125].

Figure 5. Courtesy [125] NLRP3 inflammasome priming and activation. NLRP3 inflammasome activation requires two signals: Signal 1 (initiation) is provided by activating cytokines or PAMPs, resulting in transcriptional up regulation of normative and non-standard NLRP3 inflammasome components. Signal 2 (activation) is activated by K+ efflux, Ca2+ influx, and mitochondrial reactive oxygen species, etc. ASC; IL-1R1, IL-1 receptor type1; NEK7, NIMA-related kinase7; NF-κB, nuclear factor-κB; P2X7, P2X purinoceptor7; ROS, reactive oxygen species; TLR, toll-like receptor; TNF α, tumor necrosis factor α; TNFR, tumor necrosis factor receptor; TXNIP, thioredoxin-interacting protein; MD2, myeloid differentiation factor 2;LPS, lipopolysaccharide; ER Stress, endoplasmic reticulum; TIRF, toll-interleukin 1 receptor (TIR)-inducing interferon; MyD88, myeloid differentiation primary response gene 88.

Figure 6. Courtesy [125] Diabetes Mellitus promotes the development of atherosclerosis: the role of NLRP3. In the context of diabetes mellitus, risk factors such as high glucose, oxidative stress, and inflammation can induce endothelial dysfunction, inflammation, and platelet activation and aggregation by regulating NLRP3 inflammasome, thereby promoting atherosclerosis.
Role of melatonin
Melatonin that is a pineal hormone possessing antioxidant along with anti-inflammatory characteristics have been believed to be responsible for circadian dysrhythmia-correlated with DM. Reduction in the quantities of melatonin as well as a functional association of melatonin has been illustrated with insulin that is held responsible for pathogenesis of type 2 Diabetes (T2D). In addition to those genomic studies, demonstrated that rare variation of melatonin receptor ib (MTNR1B) are further correlated with dysfunctional glucose tolerance along with escalated risk of T2D. Moreover, external melatonin treatments in cell lines, rodent models, as well as Diabetic patients have demonstrated robust actions in amelioration of Diabetes along with its associated complications. This emphasizes on the part of melatonin in glycemic homeostasis. Nevertheless, conflicting documentations exist regarding melatonin supplementation. Hence it becomes necessary to evaluate if one can translate melatonin from bench towards bedside regarding DM management. Thus Patel et al. [126], recently reviewed the therapeutic implications of melatonin treatment in different animal models as well as if it was feasible to be taken into account as a safe agent for therapy of Diabetes complications along with other Diabetes presentations like OS, ER stress, mitochondrial impairment as well as mitochondrial decontrolling, etc., (Figure 7&8) [126,127].
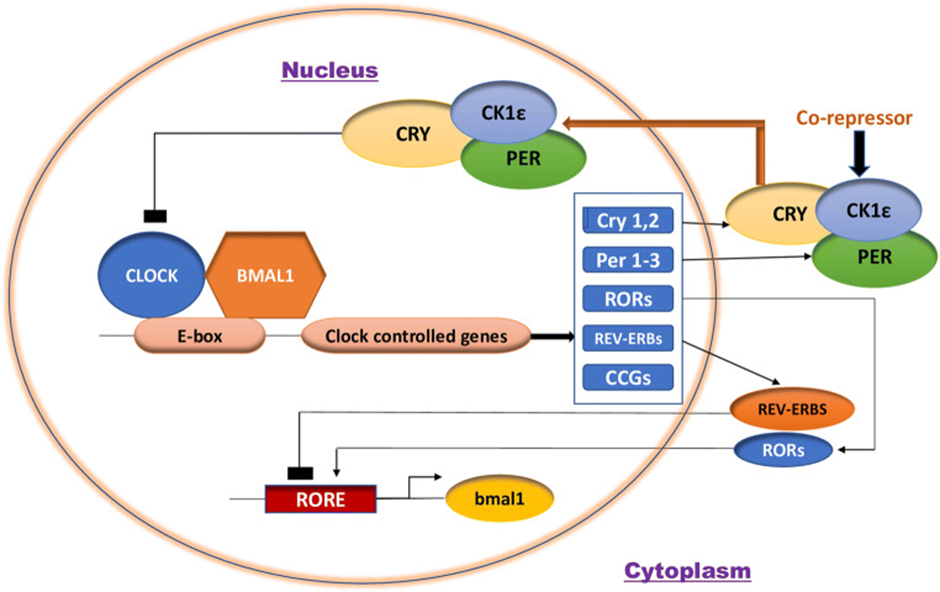
Figure 7. Courtesy [124] Molecular machinery of the Mammalian Circadian clock.

Figure 8. Courtesy [126] Circadian rhythm disruption leading to an increased susceptibility to cardiometabolic disorders.
Role of calystegines
Kowalczuk et al. [127], conducted the assessment of Calystegines in an experimental model of hyperglycemia which was triggered on human Adipose-derived stem cells (ASC). Different cellular pathways inclusive of apoptosis, Oxidative along with ER stress, inflammation along with phosphatidyl inositol 3-kinase (PI3K)/AKT protein kinase B (aliasPKB)/mTOR (mammalian target of rapamycin inhibitors) pathway metabolic correlated axis was analyzed with utilization of RT-qPCR, western block as well as flow cytometry. Treatment of HuASCs cells with Calystegines robustly facilitated the hyperglycemic cells survival along with reduced OS, failing mitochondrial dynamics besides ER stress along with enhancing the endogenous cellular antioxidant defenses. Intriguingly, nortropane alkaloids avoided the hyperglycemia-modulated inflammatory reaction, as validated by control of pro as well as anti-inflammatory reactions in HuASCs cells. Finally, they validated that Calystegines might have a protective impact on metabolic function of HuASCs cells via the rectification of aberrant AKT/PI3K/mTOR pathway. In to this study proved that Calystegines possessed the capacity of protecting HuASCs cells against hyperglycemia stimulated cellular impairment which was brought about by facilitation of AKT/PI3K/mTOR pathway [128].
Role in Diabetic Retinopathy (DR)
The generation of Diabetic Retinopathy (DR) implicates a complicated kind of pathways like Oxidative stress, inflammation, apoptosis, neurodegeneration, lipid peroxidation, angiogenesis along with ER stress with everyone of these being probable therapeutic biomarkers. A protein particular treatment has not been generated till date in view of besides glycemic regulation till the generation of proliferative stage leading to poor prognosis regarding vision for the pt. Robles-Rivera et al. [128], conducted assessment of various dietary regimens like Mediterranean diet, dietary paradigm to halt hypertension (DAHH) along with their functional foods as well as low caloric diets (LCD’s). Nutraceuticals have been evaluated in DR in view of their antioxidant, anti-inflammatory along with anti angiogenic characteristics, that might have a considerably significant impact on the pathophysiology of DR. These nutraceuticals have illustrated lesser quantities of ROS, significant inflammatory factors, cytokines as well as endothelial injury biomarkers as mono or combination therapies or simultaneously with ongoing DM therapy or nonstandard adjuvant agents like local nonsteroidal anti-inflammatory drugs (NSAIDS) (Figure 9) [129].

Figure 9. Courtesy [129], DR's physiopathology and biomarkers for each intervention. Hyperglycemia induces a variety of biochemical responses derived in angiogenesis (as shown by VEGF and endothelial damage), apoptosis, and retinal dysfunction; here, we describe where each intervention acts in these complex pathways by inhibiting the shown biomarker. Abbreviations: VEGF: vascular endothelial growth factor; G3P: glyceraldehyde 3-phosphate; DAG: diacylglycerol; AGEs: advanced glycation end products; RAGE: receptor for advanced glycation end products; PKC: protein kinase C; NOX: NADPH (nicotinamide adenine dinucleotide phosphate) oxidase; ROS: reactive oxygen species; SOD: superoxide dismutase; MnSOD: manganese superoxide dismutase; GPx: glutathione peroxidase; GR: glutathione reductase; ICAM-1: intercellular adhesion molecule-1; VCAM-1: vascular cell adhesion molecule-1; MCP-1: monocyte chemoattractant protein-1; TNF-α: tumor necrosis factor-alpha; IL-6: interleukin-6; IL-8: interleukin-8; IL-1β: interleukin-1β; PUFAs: polyunsaturated fatty acids; AIF-1: apoptosis-inducing factor-1; Bax: Bcl-2-associated X protein; LEAM: leukocyte-endothelium adhesion molecules; DASH: Dietary Pattern to Stop Hypertension.
Role of small molecule hampering agents
Furthermore, with the escalated insight agents that target these ER stress have been generated as illustrated by Krishna Prasad et al. in a recent review [130]. Assessment of a broad variety of natural along with synthetic substances by screening them regarding their characteristics of hampering ER stress in different animal models. They detailed the validating proof regarding these small molecule hampering agents as well as their pharmacological modes in amelioration of ER stress in Diabetes mellitus [130].
CONCLUSIONS
In the past 20years Comprehensive knowledge regarding molecular actors in the pathways of ER along with inflammatory stress have been acquired, with simultaneously taking part in etiopathogenesis in addition to pathophysiology of multiple diseases has been enhancing. Intriguingly, certain actors that share the 2 pathways have been enhancingly illustrated, pointing towards a complex as well as close crosstalk amongst ER along with inflammatory stress.
ER stress along with inflammatory stress are repeatedly observed to be implicated in the pathophysiology of DM regarding modes correlated with glycemia regulation (insulin liberation along with action) or in the generation/ propagation of DM complications. Moreover, hyperglycemia enhances the production of AGEs, that in turn aggravated ER stress along with inflammatory stress, aiding in the dysfunctional glucose homeostasis along with exaggeration of DM complications.
Here the molecular modes implicated in the AGEs stimulated activation of ER stress along with inflammatory stress pathways causing suppression of Slc2a4 gene which is followed by reduction in quantities of GLUT4 in AT as well as muscles. This event decreases clearance of plasma glucose, thus aiding in hyperglycemia, thus forming a vicious cycle in DM. In addition to that AGEs stimulated activation of ER stress along with inflammatory stress pathways modulate the generation of atherogenesis aiding in the propagation of DM.
Till date it has become apparent in DM regarding the dysfunctional glucose homeostasis being along with generation of neurodegenerative conditions take place secondary to AGEs stimulated ER stress along with inflammatory stress. Blockade of stimulated ER stress along with inflammatory stress under certain experimental conditions in particular tissues have shown remarkable improvement of glycemic regulation or avoidance of CVD generation or its propagation. Hence dependent on hampering of ER along with inflammatory stress greater work is the requirement regarding small molecule hampering agents acting in the form of probable targets for the avoidance of along with generation of therapies for DM or its complications. Furthermore, roles of melatonin, NLRP3, nutraceuticals are warranted in human beings for translation from bench to bedside.
REFERENCES
- Fourohi NG, Wareham NJ. (2014). Epidemiology of Diabetes. Medicine (Abingdon). 42(12):698-702.
- American Diabetes Association. (2011). Diagnosis and classification of Diabetes mellitus. Diabetes Care. 34(Suppl 1):S62-S69.
- Artime E, Romera I, Diaz-Cerezo S, Delgado E. (2021). Epidemiology and economic burden of cardiovascular disease in patients with type2 Diabetesmellitus in Spain: a systematic review. Diabetes Ther. 12(6):1631-1659.
- Lester E. (1989). The Clinical value of glycated haemoglobin and glycated plasmaproteins. Ann Clin Biochem. 26(Pt 3):213-219.
- Pinto Junior DC, Pinto SilvaKS, Fabre NT Michalani ML, Yonamine CY, Esteves JV, Fabre NT, et al. (2018). Advanced Glycation end-products-induced insulin resistance involves skeletal muscles GLUT4 expression. Sci Rep. 8:8109.
- Pinto RS, Machado UF, Passarelli M. (2020). Advanced Glycation end-products as biomarkers for cardiovascular disease: browning clarifying atherogenesis. Biomark Med. 14(8):611-614.
- Kaur KK, Allahbadia GN, Singh M. (2021). Role of Adipocyte impairment in Heart Failure Induction in subjects that are obese along with prediabetes and overt Diabetes mellitus-A Systematic Review. J Cardiol &Card Disord. 2(1):1-21.
- Kaur KK,Allahbadia GN, Singh M. (2022). An Update on the Risk Factors Correlating NAFLD with Cardiovascular Disease: Specifically Mitochondrial- Fatty Acids β Oxidation in Liver with Therapeutic Approaches of Avoidance of CVD Associated Mortality-ASystematic Review. J Endocrinol. 6(1):1-20.
- Kaur KK, Allahbadia GN, Singh M. (2021). Potential role of Epigenetic Modulation in prevention or therapy for Diabetic Kidney Disease-still a dream or a reality-A Systematic Review. J Diab Nephro Diab Mgmt. 1(1):1-26.
- Kaur KK, Allahbadia GN, Singh M. (2022). Diabetic Cardiomyopathy: An Update on its Pathophysiology with Specific Emphasis on Epigenetics Modifications Besides Treatment-A systematic review. BIJRDPM. 1( 1):1-16.
- Kaur KK, Allahbadia GN, Singh M. (2021). The utility of phytochemicals obtained from plants for the treatment of type 2 Diabetes Mellitus with Emphasis on the Epigenetic Alterations related to T2DM & their Impact as Therapeutic Agents in the form of so called Epi-drugs: a systematic review. Adv Obes Weight Manag Control. 11(6):195-206.
- Kaur KK, Allahbadia GN, Singh M. (2019). Importance of Simultaneous Treatment of Obesity and Diabetes Mellitus: A Sequelae to the Understanding of Diabesity-A Review. Obes Res Open J. 6(1):1-10.
- Kaur KK, Allahbadia GN, Singh M. (2021). Role of Trigonella foenum-graecim Extract along with Ursolic Acid a Pentacyclic Triterpenoid as Newer Plant Productsfor the Therapy of Diabetes Mellitus-A Short Communication. Acta Scientific Nutritional Health. 5(6):12-15.
- Kaur KK, Allahbadia GN, Singh M. (2019). Development of protein tyrosine phosphatase 1B (PTPIB) Inhibitors from marine sources and other natural products-Future of Antidiabetic Therapy: A Systematic Review. Korean Journal of Food & Health Convergence. 5(3):21-33.
- Kaur KK, Allahbadia GN, Singh M. (2022). An Update on the Therapeutic Potential of Herbal Preparations with regards to Molecular & Biochemical Mechanisms in the Management of Diabetes Mellitus: a Systematic Review. World Journal of Advance Healthcare Research. 6(3):1-17.
- Kaur KK, Allahbadia GN, Singh M. (2020). Utilization of extracellular vesicles for treatment of Type 1 Diabetes Mellitus (T1DM) along with 2 T2DM besides Complications associated with Diabetes-A Systematic Review. J Clin Diabetes Obes. 1:001-013.
- Kaur KK, Allahbadia GN, Singh M. (2021). Would it be advantageous utilizing beta cell therapies over immunotherapies for avoidance of Type 1 Diabetes-A Systematic Review on the role of beta cells in etiopathogenesis of Type 1 Diabetes along with treatments targeting beta cells or combination therapy would be better. Aditum Journal of Clinical and Biomedical Research. 1(2).
- Kaur KK, Allahbadia GN, Singh M. Are we Any Close to Unraveling the Mechanism of Interactions Among Susceptibility Genes Towards Type 1 Diabetes, Gut Microbiota Along with Environmental Factors, Specifically Early Diet Patterns-A Systematic Review. Journal of Endocrinology and Surgical Endocrinology. 2(1):1-20.
- Kaur KK, Allahbadia GN, Singh M. (2021). Targeting macrophage polarization for therapy of diabesity–the feasibility of early improvement of insulin sensitivity and insulinresistance-a comprehensive systematic review. J Diab Metab Disorder Control. 8(1):6-25.
- Kaur KK, Allahbadia GN, Singh M. (2021). How can we Use Empagliflozin as an Adjuvant in Reducing Required Need of Insulin in Type 1 Diabetes along with Lowered HbA1c, Weight without Fear of DKA-A Mini Review. J Clin Cases Rep. 4(2):30-38.
- Kaur KK, Allahbadia GN, Singh M. (2019). Advantage of Cardiovascular Outcome Trials (CVOT’s) for SGLT2 (Sodium Glucose Transporter 2) Inhibitors in Type 2 Diabetes Mellitus (T2 DM). EC Endocrinology and Metabolic Research. 4(9):38-44.
- Klip A, McGraw TE, James DE. (2019). Thirty sweet years of GLUT4. J Biol Chem. 294(30):11369-11381.
- Frendo Cumbo S, Tokarz VL, Bilan PJ, Brumell JH, Klip A. (2021). Communication between autophagy and insulin action-insulin resistance? Front Cell Dev Biol. 9:708431.
- Sajan MP, Lee MC, Foufelle F, Sajan J, Clelaland L, Farese RV. (2018). Coordinated regulationof hepatic FoxO1,PGC-1-α and SREBP1c facilitates insulin action and resistance. Cell Signal Med.43:62-70.
- James DE, Strube M, Mueckler M. (1989). Molecular cloning ion characterization of an insulin regulatable promoted glucose transporter. Nature. 338(6210):83-87.
- Salazar MR, Carbajal HA, Espeche WG, Aizpurua M, Leiva Sisniguez CE, Leiva Sisniguez BC, et al. (2016). Insulin resistance: the lynchpin between prediabetes and cardiovascular disease. DiabVasc Dis Res. 13(2):157-163.
- Tirosch A, Tuncman G, Calay ES, RathausM, Ron L, Tirosch A, et al. (2021). Intercellular transmission of hepatic ER stress in obesity disrupts systemic metabolism. Cell Metab. 33(2):319-333.
- Mueckler M, Thorens B. (2013). The SLC (GLUT4) family of membrane transporters. Mol Asp Med. 34(2-3):121-138.
- Karnieli E, Armoni M. (2008). Transcriptional regulation of the insulin responsive glucose transporter GLUT4 gene: from physiology to pathology. Am J Physiol Endocrinol Metab. 295(1):E38-E45.
- Machado UF, Shimizu I, Saito M. (1994). Reduced content and preserved translocation of glucose transporter( GLUT4)in White Adipose tissue of obese mice. Physiol Behav.55:621-625.
- Zisman A, Peroni OD, Abel ED, Michael MD, Mauvais Jarvis F, Lowell BB, et al. (2000). Targeted disruption of the glucose transporter 4 selectively in muscle causes insulin resistance and glucose intolerance. Nat Med.6(8):924-928.
- Einarson IR, Acs A, Ludwig C, Panton UH. (2018). Prevalence of cardiovascular disease in type2 Diabetes:a systematic literature review of scientificevidence from across the world in 2007-2017. Cardiovasc Diabetol. 17:83.
- Eckel RH, Bornfeldt KE, Goldberg IJ. (2021). Cardiovascular disease in Diabetes beyond glucose. Cell Metab. 33(8):1519-1545.
- Thambiah SC, Lai LC. (2021). Diabetic dyslipidemia. Pract Lab Med. 26:e00248.
- Giacco F, Brownlee M. (2010). Oxidative stress and Diabetic complications. Circ Res. 107(9):1058-1070.
- Myoishi M, Hao H, Minamino T, Watanabe K, Nishihira K, Hatakeyama K, et al. (2007). Increased endoplasmic reticulum stress in atherosclerotic plaques associated with acute coronary syndrome. Circulation. 116(11):1226-1233.
- Morgenstern J, Campos Campos M, Nawroth P, Fleming P. (2020). The glyoxalase system-new insights into an ancient metabolism. Antioxidants. 9(10):939.
- Sell DR, Sun W, Gao X, Straunch C, Lachin JM, Cleary P, Genuth S, DCCT/EDIC Research Group, Monnier VM. (2016). Skin collagen fluorophores LW-1versus Skin fluorescence as markers for the long term progression of macrovascular disease in type1 Diabetes. Cardiovasc Diabetol. 15:30.
- Genuth S, Sun W, Cleary P, GaoX, Sell DR, Lachin JM. DCCT/EDIC Research Group, Monnier VM. (2015). Skin advanced glycation end-products glucosepane, methylglyoxal hydroimidazolones are independently associated with long term microvascular complications progression of type1 Diabetes. Diabetes. 64(1):266-278.
- Akirav EM, Preston Helburt P, Garyu J, Hanegariu O, Clynes R, Schmidt HM, et al. (2012). RAGE expression in human T cells: a link betweenenvironmentfactors and adaptive immune responses. PLoS ONE. 7(4):e34698.
- Prasad K. (2014). Low levels of serum soluble receptors advanced glycation end-products, biomarkers for diseased state: myth or reality. Int J Angiol. 23(1):11-16.
- Santos-Bezerra DP, Machado-Lima A, Monteiro MB, Admoni SN, Perez RV, Machado CG, et al. (2018). Dietary advanced glycation end-products and medicines expression of SIRT1 and DDOSTin in peripheral blood mononuclear cells from long term type1 Diabetic patients. DiabVasc Dis Res. 15(1):81-89.
- Rabbani N, Godfrey L, Xue M, Shahe F, Geoffrion M, Milne R, et al. (2011). Glycation of LDL by methylglyoxal increases arterial atherogenicity: a possible contributor to increased risk of cardiovascular disease in Diabetes. Diabetes. 60(7):1973-1980.
- Godfrey L, Yamada-Fowler N, Smith J, Thornalley PJ, Rabbani N. (2014). Arginine directed glycation and decreased HDL plasma Concentration in muscle and functionality. Nutr Diabetes. 4(9):e134.
- Leung PS, Gershwin ME. (1991). The immuno biology of heat shockproteins. J Investigol Allergol Clin Immunol. 1:13-20.
- Schroder M, Kaufman RJ. (2005). ER stress and unfolded protein response. Mutat Res. 569(1-2):29-63.
- Hardinge HP, Zeng H, Zhang Y, Jungries R, Chung P, Plesken H, et al. (2001). Diabetes mellitus and exocrine pancreatic dysfunction in perk-/-micereveals a role for translational control in secretory cell survival. Mol Cell. 7(6):1153-1163.
- Iwawaki T, Akai R, Kohno K. (2010). IRE1a disruption causes histological abnormality of exocrine tissues, increase of blood glucose level and decrease of serum immunoglobulin level. PLoS ONE. 5(9):e13052.
- Oyadomari M, Koizumi A, Takeda K, Gotoh T, Akira S, Araki E, et al. (2002). Targeted disruption of the Chop gene delays endoplasmic reticulum stress-mediated Diabetes. J Clin Investig. 109(4):525-532.
- Ye R, Jung DY, Jun JY, Li J, LuoS, Ko HJ, et al. (2010). Grp78 heterozygosity promotes adaptive unfolded protein response and attenuates diet induced obesity and insulin resistance. Diabetes. 59(1):6-16.
- Walters ST, Luzuriaga J, Chan JY, Grey ST, Laybutt DR. (2013). Influenceof chronic hyperglycemia on loss of adaptive unfolded protein response in transplanted islets.J Mol Endocrinol. 51(2):225-232.
- Pandey VK, Mathur A, Kakkar P. (2019). Emerging role of unfolded protein response (UPR) mediated proteotoxic apoptosis in Diabetes. Life Sci. 216:246-258.
- Mukherjee N, Lin L, Contreras CJ, Templin AT. (2021). B cell death and Diabetes: past discoveries,present understanding and potentialfuture advances. Metabolites. 11(11):796.
- Hotamisligil GS. (2006). Inflammation and metabolic disorders. Nature. 444(7121):860-867.
- Ruan H, Hacohen N, Golub TR, Van Parijs L, Lodish HF. (2002). Tumor necrosis factor alpha suppresses adipocyte specific genes and activates expression ofpreadipocyte genes in 3T3L1 adipocytes: nuclear factor κB activation by TNF-alpha is obligatory. Diabetes. 51(5):1319-1336.
- Tan AB, Mercado EL, Hoffmann A, Niwa M. (2012). ER stress activates NFκB integrating functions of basal IKKactivity, IRE1 and PERK. PLoS ONE. 7(10):e45078.
- Fernandez D, Geisse A, Bernales JL, Lira A, Osorio F. (2021). The unfolded protein response in immune cells as an emerging regulator of neuroinflammation. Front Aging Neurosci. 13:682633.
- Fatima LA, Campello RS, Barreto-Andrade JN, Passarelli M, Santos RS, Clegg DJ, Machado UF. (2012). Estradiol stimulates adipogenesis and Slc2a4/ GLUT4 expression via ESR-1 mediated activation of CEBPA. Mol Cell Endocrinol. 498:110447.
- Jang EH, Ko JH, Ahn CW, Lee HH, Shin JK, Chang SJ, et al. (2010). In vivo and in vitro application of black soybean peptides in theamelioration of endoplasmic reticulum stress and improvementof insulin resistance. Life Sci. 86(7-8):267-274.
- Raciti GA, Iadicicco C, Ulianich L, Vind BF, Gaster M, Andreozzi F, et al. (2010). Glucosamine induced endoplasmic reticulum stress affects GLUT4 expression via activating transcription factor6 in rat and human skeletal muscle cells. Diabetologia. 53(5):955-965.
- SilvaJ L, Giannocco G, Furuya DT, Lima GA, Moraes PA, Nachef S, et al. (2005). NF-kappaB, MEF2A, MEF2D and HIF1-a involvement on insulin and contraction induced regulation of GLUT4 gene expression in soleus muscle. Mol Cell Endocrinol. 240(1-2):82-93.
- Furuya DT, Poletto AC, Favarro RR, Martins JO, ZornTM, Machado UF. (2010). Antiinflammatory effect of atorvastatin ameliorates insulin resistance in monosodium glutamate treated mice. Metabolism. 59(3):395-399.
- Furuya DT, Poletto AC, Silva Esteves JV, Freitas HS, Machado UF. (2012). Inhibition of Cannabinoid receptor1 (CBR1) upregulates Slc2a4 expression via nuclear factor kappaB and sterol regulatory element binding protein 1c in adipocytes. J Mol Endocrinol. 49(2):97-106.
- Moraes PA, Pinto Junior DC, Esteves JVDC, Yonamine CY, Machado UF, Mori RC. (2014). Insulin acutely triggers transcription of Slc2a4 gene: participationof the AT-rich,E-box and NFκB binding sites. Life Sci. 114(1):36-44.
- Poletto AC, Furuya DT, David-Silva A, Ebersbach-Silva P, Santos RS, Correa-Giannela ML, et al. (2015). Oleic acid and linoleic acid fatty acids downregulate Slc2a4/ GLUT4 expression via NFκB and SREBP1c in skeletal muscle cells. Mol Cell Endocrinol. 401:65-72.
- Ebersbach-Silva P, Poletto AC, David-Silva A, Seraphim PM, Anhe GF, Passarelli M, Furuya DT, Machado UF. (2018). Palmitate induced Slc2a4/GLUT4 downregulation in L6 muscle cells:evidence of inflammatory and endoplasmic reticulum stress involvement. Lipids Health Dis. 17:64.
- Liong S, Lappas M. (2016). Endoplasmic reticulum stress regulates inflammation and insulin resistance in skeletal muscles from pregnant women. Mol Cell Endocrinol. 425:11- 25.
- Lin MW, Chen CL, Cheng TT, Huang CC, Tsai JW, Feng GM, et al. (2021). Prolonged preoperative fasting induces postoperative insulin resistance by ER stress mediated Glut4 downregulation in skeletal muscles. Int J Mol Sci. 18(5):1189-1197.
- Obafemi TO, Olasehinde OR, Olaoye OA, JaiyesimiKF, Adefum FD, Adewale OB, et al. (2020). Metformin/donezepil combination modulates brain antioxidant status and hippocampal endoplasmic reticulum stress in type2 Diabetic rats. J Diabetes Metab Disord. 19(1):499-510.
- Varghese DS, Ali BR. (2021). Pathological Crosstalk Between Oxidized LDL and ER Stress in Human Diseases: A Comprehensive Review. Front Cell Dev Biol. 9:674103.
- Hong D, Bai YP, Gao HC, Wang X, Li LF, Zhang GG, et al. (2014). Ox-LDL induces endothelial cells apoptosis via the LOX1 dependent endoplasmic reticulum stress pathway. Atherosclerosis. 235(2):310-317.
- Tsukano H, Gotoh T, Endo M, Miyata K, Tazume H, Kadomatsu T, et al. (2010). The endoplasmic reticulum stress- C/EBP-homologous protein mediated apoptosisin macrophages undergoing endoplasmic reticulum stress contributes to the instability of atheroscleroticplaques. Arterioscler Thromb Vasc Biol. 30(10):1925-1932.
- Ren J, Bi Y, Sowers JR, Hetz C, Zhang Y. (2021). Endoplasmic reticulum stress and unfolded protein response in cardiovascular disease. Nat Rev Cardiol. 18(7):499-451.
- Zhou Y, Murugan DD, Khan H, Huang Y, Cheang WS. (2021). Roles and therapeutic implicationsof endoplasmic reticulum stress and Oxidative stress in cardiovascular diseases. Antioxidants. 10(8):1167.
- Zhou Y, Wan X, Seidel K, Zhang M, Goodman JB, Seta F, et al. (2021). Hypercholesterolemia differentially affects the unfolded protein response in the vasculature of Apoe-/- mice. J Am Heart Assoc. 10(18):e020441.
- Beriault DR, Sharma S, Shi Y, Khan MI, Westuck GH. (2011). Glucosamine supplementation promotes endoplasmic reticulum stress, hepatic steatosis and accelerated atherogenesis in Apoe-/- mice. Atherosclerosis. 219(1):310-317.
- Chong WC, Shastri MD, Peterson GM, Patel RP, Pathinayake PS, Dua K, et al. (2021). The complex interplay between endoplasmic reticulum stress and NLRP3 inflammasome: a potential therapeutic target for inflammatory disorders. Clin Transl Immunol. 10(2):e1247.
- Zeng L, Lu M, Mori K, Luo S, Lee AS, ZhuY, et al. (2004). ATF6 modulates SREBP2-mediated lipogenesis. EMBO J. 23(4):950-958.
- Xu X, Lei T, Li W, Oh H. (2019). Enhanced cellular cholesterol efflux by naringin is mediated through inhibiting endoplasmic reticulum stress - ATF6 activity in macrophages. Biochim Biophys Acta Mol Cell Biol Lipids. 1864(10):1472-1482.
- Che X, Xiao Q, Song W, Zhang H, Sun B, Geng N, et al. (2021). Protective function of Liver X receptor-α expression in hepatocytesin established vulnerable plaques involvement of regulating endoplasmic reticulum-mediated macrophage apoptosis efferocytosis. J Am Heart Assoc. 10(10):e018455.
- Song G, Wu X, Zhang P, Yu Y, Yang M, Jiao P, et al. (2016). High density lipoprotein inhibits oxLDL inducedadipokine secretion by upregulating SR-B1 expression and suppressing ER stress pathway. Sci Rep. 6:30889.
- Navas Madronal M,Castelblanco E, Camacho M, Consegal M, Ramirez-Morros A, Sarrias MR, et al. (2020). Role of the scavenger receptor CD36 in accelerated Diabetic atherosclerosis. Int J Mol Sci. 21(19):7360.
- Rabbani N, Xue M, Thornalley PJ. (2021). Dicarbonyl stress, protein glycation and the unfolded protein response. Glycoconj J. 38(3):331-340.
- Frederickson EK, Rosenbaum JC, Locke MN, Milac TL, Gardner RG. (2011). Exposed hydrophobicity is a key determinant of nuclear quality and control degradation. Mol Biol Cell. 22(13):2384-2395.
- Saha K, Kim SJ, Zhang Z, Lee YC, Sarkar C, Tsai PC, et al. (2008). RAGE signaling contributes to neuroinflammation in infantile neuronal ceroid lipofuscinosis. FEBS Lett. 582(27):3823-3831.
- Irshad Z, Xue M, Ashour A, Larkin JR, Thornalley PJ, Rabbani N. (2019). Activation of the unfolded protein response in high glucose treated endothelial cells is mediated by glyoxal. Sci Rep. 9(1):7889.
- Nourbakhsh M, Sharifi R, Heydari N, Nourbakhsh M, Ezzati-Moubaser S, Zarrinahad H. (2022). Circulating TRB3 and GPR78 levels in type2 Diabetic patients: between glucose homeostasis and endoplasmic reticulum stress. J Endocrinol Investig. 45(3):649-655.
- Garay Sevilla ME, Rojas A, Porteo Otin M, Uribarri J. (2021). Dietary AGEs asexogenous boosters of inflammation. Nutrients. 13(8):2802.
- Uribarri J, Cai W, Pyzik R, Goodman S, Chen X, Zhu L, et al. (2014). Suppression of native defense mechanisms, SIRT1 PPARγ, by dietary glycooxidants precedes disease in adult humans, relevance to lifestyle -engendered chronic diseases. Amino Acids. 46(2):301-309.
- Mondel M, Koyama H, Otsuka Y, Morioka T, Mori K, Shoji T, et al. (2013). Receptor for advanced glycation end-products regulates adipocyte hypertrophy and insulin sensitivity in mice: involvement of toll like receptor 2. Diabetes. 62(2):478-489.
- Gaens KHJ, Goossens GH, Niessen PM, Van Greevenbroek MM, Van Der Kallen CJH, Niessen HW, et al. (2014). Nε (carboxymethyl) lysine receptor for advanced glycation end-products axisis a key modulator of obesity induced dysregulation of adipokine expression and insulin resistance. Arterioscler Thromb Vasc Biol. 34(6):1199-1208.
- Chilelli NC, Faggian A, Favaretto F, Milan G, Compagnin C, Dassie F, et al. (2021). In vitro chronic glycation induces AGEs accumulation reducing insulin stimulated glucose uptake and increasing GLP1R in adipocytes. Am J Physiol Endocrinol Metab. 320(5):E976-E988.
- Dozio E, Vianello E, Briganti S, Lamont J, Tacchini L, Schmitz G, et al. (2016). Expression of the receptor for advanced glycation end-products in epicardial fat: link with tissue thickness and local insulin resistance in coronary artery disease. J Diabetes Res. 2016:2327341.
- DharA, Dhar L, Jiang B, Desai KM, Wu L. (2011). Chronic methylglyoxal infusion by minipump causes pancreatic beta cell dysfunction and induces type2 Diabeteseveals in Sprague Dawley rats. Diabetes. 60(3):899-908.
- Da Silva KS, Pinto PR, Fabre NT, Gomes DJ, Thieme K, Okuda LS, et al. (2017). N-acetylcysteine counteracts Adipose tissue macrophage infiltration and insulin resistance elicited by advanced glycated albumin in healthy rats. Front Physiol. 8:723.
- Passarelli M, Machado UF. (2022). AGEs-induced and Endoplasmic reticulum stress/inflammation-mediated regulation of GLUT4 expression and atherogenesis in Diabetes mellitus. Cells. 11(1):104.
- Okuda LS, Castilho G, Rocco D, Nakandare ER, Catanozi S, Passarelli M. (2012). Advanced glycated albumin impairs HDL anti-inflammatory activity and primes macrophages for inflammatory responses that reduces Reverse cholesterol transport. Biochim Biophys Acta. 1821(12):1485-1492.
- Iborra RT, Machado-Lima A, Castilho G, Nunes VS, Abdalla DS, Nakandare ER, Passarelli M. (2011). Advanced glycation in macrophages induces intracellular accumulation of 7 ketocholesterol and total sterols by decreasing the expression of ABCA1 and ABCG1. Lipids Health Dis. 10: 172.
- Gomes DJ, Velosa A, Okuda LS, Fusco FB, da Silva KS, Pinto PR, et al. (2016). Glycated albumin induces lipid infiltration in mice aorta independent of DM and RAS local modulation by inducing lipid peroxidation and inflammation. J Diabetes Complications. 30(8):1614-1621.
- De Souza PR, Castilho G, Paim BA, Machado-Lima A, Inada NM, Nakandare ER, et al. (2012). Inhibition of macrophage Oxidative stress prevents the reduction of ABCA1 transporter induced by advanced glycated albumin. Lipids. 47(5):443-450.
- Castilho G, Okuda LS, Pinto RS, Iborra RT, Nakandare ER, Santos CX, et al. (2012). ER stress is associated with reduced ABCA1 in macrophages treated with advanced glycated albumin. reversal by a chemical chaperone. Int J Biochem Cell Biol. 44(7):1078-1086.
- Mizuno T, Hayashi H, Naoi S, Sugiyama Y. (2011). Ubiquitination is associated with lysosomal degradationof cell surface-resident ATP binding cassette transporter A(ABCA1)through the endosomal sorting complex required for transport (ESCRT) pathway. Hepatology. 54(2): 631-643.
- Iborra RT, Machado-Lima A, Okuda LS, Pinto PR, Nakandare ER, Machado UF, et al. (2018). AGE-albumin enhances ABCA1degradation by ubiquitin-proteasome and lysosomal pathways in macrophages. J Diabetes Complications. 32(1):1-10
- Minnani C, Machado-Lima A, Iborra RT, Okuda LS, deSouza PR, Santana MFM, et al. (2021). Persistent effects of advanced glycated albumin driving inflammation and disturbances in cholesterol efflux in macrophages. Nutrients. 13(10):3633.
- Machado-Lima A, Lopez-Diez R, Iborra RT, Pinto RS, Daffu G, Shen X, Nakandare ER, Machado UF. (2020). RAGE mediates cholesterol efflux impairment in macrophages caused by human advanced glycated albumin. Int J Mol Sci. 21(19):7265.
- Machado JT, Iborra RT, Fusco FB, Castilho G, Machado-Lima A, Nakandare ER, et al. (2014). N-acetylcysteine prevents endoplasmic reticulum stress in macrophages by serum albumin drawn from Chronic Kidney Disease rats and selectively affects lipid transporters, ABCA1 and ABCG1. Atherosclerosis. 237(1):343-352.
- De Araujo Lira AL, de Fatima Mello Santana M, de Souza PR, Minnani C, Iborra RT, de Lima AMS, et al. (2021). Serum albumin modified by carbamoylation impairsmacrophage cholesterol efflux in Diabetic Kidney Disease. J Diabetes Complications. 35(9):107969.
- Chen Y, Liu CP, X KF, Mao XD, Lu YB, Fang L, et al. (2008). Effect of taurine conjugated ursodeoxycholic acid on endoplasmic reticulum stress and apoptosis induced by advanced glycation end-products in cultured mouse podocytes. Am J Nephrol. 28(6):1014-1022.
- Jeong SR, Lee KW. (2021). Methylglyoxal-derived advanced glycation end- products (AGE4) induced apoptosis leads to mitochondrial dysfunction and endoplasmic reticulum stress through the RAGE/JNK pathway in kidney cells. Int J Mol Sci. 22(12):6530.
- Liu Z, Zhang Y, Pan S, Qiu C, Jia H, Wang Y, et al. (2021). Activation of RAGE dependent endoplasmic reticulum stress reveals associates with exacerbated post myocardial infarction ventricular arrythmias in Diabetes. Am J Physiol Endocrinol Metab. 320(3):E539-E550.
- Liu Z, Zhu H, Ma Y, Tang Z, Zhao N, Wang Y, Pan S. (2021). AGEs exacerbate coronary microvascular dysfunction in NoCAD by activating endoplasmic reticulum stress- mediated PERK signaling pathway. Metabolism. 117:154710.
- Dong Y, Zhang M, Wang S, Lang B, Zhao Z, Liu C, et al. (2010). Activation of AMP-activated protein kinase inhibits oxidized LDL-triggered LDL in vivo. Diabetes. 59(6):1386-1396.
- Ravi R, Ragavachetty Nagaraj N, Subramaniam Rajesh B. (2020). Effect of advanced glycation end-products on paroxonase 2Lits impact on endoplasmic reticulum stress and Inflammation in HUVEC cells. Life Sci. 246:117397.
- Guttierez-Mascal FM, Cardelo MP, dela Cruz S, Alcala Diaz JF, Rancos RamosI, Guler L, et al. (2021). Advanced glycation end-products by Mediterranean diets is associated with increased likelihood of type2 Diabetes remission in patients with coronary heart disease:From the Cardioprev study. MolNutrFood Res. 65(1):e1901290.
- Koksal AR, Vernel GN, Zhou Q. (2021). Endoplasmic reticulum stress in biological processing and disease. J Investig Med. 69(2):309-315.
- David-Silva A, Esteves JV, Moraes MRPT, Freitas HS, Zorn TM, Correa-Giannela ML, et al. (2020). Dual SGLT1/ SGLT2 inhibitor phlorizin ameliorates Non alcoholic fatty liver disease and hepatic glucose production in type2 Diabetic mice. Diabetes Metab Syndr Obes. 13:739-751.
- Pavlovic N, Kopsida M, Gerwins P, Heindryckyckz F, et al. (2020). Inhibiting P2Y12 in Macrophages Induces Endoplasmic Reticulum Stress and Promotes an Anti-Tumoral Phenotype. Int J Mol Sci. 21(21):8177.
- Mastrocola R, Nigro D, Chiazza F, Medana C, Del Bello F, BoccuzziG, et al. (2016). Fructose-derived advanced glycation end-products drive lipogenesis and skeletal muscles reprogramming via SREBP1c dysregulation in mice. Free Radical Biol Med. 91:224-235.
- Takeuchi M, Sakasai Sakai A, Takata T, Takino JJ, Koriyama Y, Kikuchi C, et al. (2021). Intracellular toxicAGEs (TAGEs) triggers numerous types of cell damage. Biomolecules. 11(3):387.
- Litwinowicz K, Wasczuk E, Gamian A. (2021). Advanced glycation end-products in common non infectious Liver diseases: systematic review and meta-analysis. Nutrients. 13(10):3370.
- Pereira ENGDS, Paula DP, de Araujo BP, de Fonseca MJM, Diniz MFHS, Daliry A, et al. (2021). Advanced glycation end-products:a potential biomarker for stratication of Non alcoholic fatty liver disease in ELSA-Brasil study. World J Gastroenterol. 27(29):4913-4928.
- Ajoolabady A, Liu S, Klionsky DJ, Lip GYH, Tuoileho J, Kavalakatt S, et al. (2021). ER stress in obesity pathogenesis and management. Trends Pharmacol Sci. 43(2):97-109.
- Kaur KK, Allahbadia GN, Singh M. (2022). An Update on the Innovative Part of NLRP3 Inflammasome Regarding Newer Strategies for Treatment of Reproductive Conditions Possessing Greater Risk: A Systematic Review. J Gynecol. 7(3):1-12.
- Zhao H, Liu H, Yang Y, Wang H. (2022). The role of H2S regulating NLRP3 inflammasome. Int J Mol Sci. 23(9):4818
- Ye JX, L Li F, Wang M, Ma T, Tian Y, Zhang Q, et al. (2022). Diabetes mellitus promotes thedevelopment of atherosclerosis:the role of NLRP3 inflammasome. Front Immunol. 13:900254.
- Wajid F, Poolacherla R, Mim FK, Bangash A, Rutkofsky IH. (2020). Therapeutic potential of melatonin asa chronobiotic and cytoprotective agent in Diabetes mellitus. J Diabetes Metab Disord. 19(2):1797-1785.
- Patel R, ParmarN,, Pramanik Palit S, Rathwa N, Ramachandran AV, Begum R. (2022). Diabetes mellitus and melatonin: Where are we? Biochimie. 202:2-14.
- Kowalczuk A, Bourebaba N, Panchuk J, Marycz K, Bourebaba L. (2022). Calystegines involves metabolic activity of human Adipose-derived stem cells(ASC) under hyperglycemic conditions reduction of Oxidative/ER stress and inflammation, and the promotion of AKT / PI3K /mTOR pathway. Biomolecules. 12(3):460.
- Robles-Rivera RR, Castellanos-González JA, Montaño CO, Flores-Martin RA, López-Contreras AK, Arevalo-Simental DE, et al. (2020). Adjuvant Diabetic retinopathy as an early approach todelay its progression: the role of Oxidative stress and inflammation. Oxid Med CellLongev. 2020:6190609.
- Krishna Prasad M, Mohandas S, Ramkumar KM. (2022). Role of ER stress inhibitors in the management of Diabetes. Eur J Pharmacol. 922:174893.