Previous Issues Volume 2, Issue 1 - 2018
Plectin, A Cytolinker Protein, Plays an Important Role in Differentiation and Actin Ring Formation in Osteoclasts
Takuma Matsubara1 , Masayuki Kinbara2 , Toshihiro Maeda2 , Mitsuhiro Yoshizawa2 , Shoichiro Kokabu1 , Teruko Takano-Yamamoto2
1Division of Molecular Signaling and Biochemistry, Department of Health Improvement, Kyushu Dental University, 2-6-1, Manazuru, Kokurakita-ku, Kitakyushu, Fukuoka, Japan. 2Dentofacial Orthopedics, Graduate School of Dentistry, Tohoku University, Sendai, Japan. Corresponding Author: Takuma Matsubara, Division of Molecular Signaling and Biochemistry, Department of Health Improvement, Kyushu Dental University, 2-6-1, Manazuru, Kokurakita-ku, Kitakyushu, Fukuoka, Japan, Tel: +233500016591 E-Mail: [email protected]
Received Date: 26 Dec 2017 Accepted Date: 22 Jan 2018 Published Date: 26 Jan 2018
Copyright © 2018 Matsubara T
Citation: Matsubara T, Kinbara M, Maeda T, Yoshizawa M, et al. (2018). Plectin, A Cytolinker Protein, Plays an Important Role in Differentiation and Actin Ring Formation in Osteoclasts. Mathews J Cytol Histol. 1(1): 006.
ABSTRACT
Osteoclasts resorb the bone matrix and maintain bone and calcium homeostasis. During bone resorption, osteoclasts form an actin ring and seal the bone matrix for bone resorption. The tyrosine kinase Src is essential for actin ring formation and bone resorption. However, the molecular mechanisms underlying the regulation of actin ring formation by Src is still unclear. A cytolinker protein, plectin, was identified as one of the proteins downstream of Src. Plectin is localized, along with Src, around the actin ring of osteoclasts. Plectin binds to and is phosphorylated by Src. Differentiation and actin ring formation were inhibited by plectin downregulation. These results suggest an important role for plectin in osteoclast differentiation and actin ring formation via phosphorylation by Src.
KEYWORDS
Osteoclast; Bone Resorption; Actin Organization; Src; Plectin.
ABBREVIATIONS
Src:Rous sarcoma oncogene; M-CSF: Macrophage colony stimulating factor; RANKL: Receptor activator NF-κB ligand; RANK:Receptor activator NF-κB; MAPK: Mitogen-activated protein kinase; NFATc1: Nuclear factor of activated T cells, cytoplasmic, calcineur in dependent 1; Pyk2: Proline-rich tyrosine kinase 2; p130Cas: Crk-associated substrate; qPCR: Quantitative polymerase chain reaction; PKC: Protein kinase C; JNK: c-Jun N-terminal kinase; Erk: Extracellular regulated MAP kinase.
INTRODUCTION
Osteoclasts are multinuclear cells differentiated from hematopoietic cells. Hematopoietic stem cells are differentiated into monocytes by the macrophage colony stimulating factor (M-CSF), which is secreted by osteoblasts or osteocytes. Subsequently, the monocytes fuse to each other and acquire osteoclast phenotypes, due to receptor activator NF-κB ligand (RANKL) and receptor activator NF-κB (RANK) signaling in the presence of M-CSF [1,2]. RANK-RANKL signaling activates NF- κB and mitogen-activated protein kinase (MAPK) signaling, such as c-Jun and c-fos, and then, promotes expression and activation of the nuclear factor of activated T cells, cytoplasmic, calcineur in-dependent 1 (NFATc1), a master regulator of osteoclast differentiation [1-3]. After differentiation, osteoclasts attach to the bone matrix through αvβ3 integrins and organize the actin cytoskeleton [4, 5]. To cover and seal the bone matrix, osteoclasts form a unique actin structure called a podosome [4, 5]. The podosome is a dot-like structure, consisting of accumulated actin, integrins, and some other actin-binding and -regulatory proteins [4]. Podosomes are arranged at the cell periphery and form a ring-like structure called the actin ring [5]. Formation of the actin ring is essential for the formation of the sealing zone and bone resorp- tion by osteoclasts [4, 6, 7]. As tyrosine kinase-deficient osteoclasts cannot form an actin ring and sealing zone, severe osteopetrosis is observed in Srcdeficient mice [6-8]. This suggests that Src regulates actin ring formation, and subsequently, bone resorption. Although Src does not contain an actin-binding domain, it regulates actin organization by binding and phosphorylating actin regulatory proteins, such as the proline-rich tyrosine kinase 2 (Pyk2) and Crk-associated substrate (p130Cas) [9-14]. The phenotypic characteristics of Pyk2-deficient mice and osteoclast-specific p130Cas-deficient mice include mild osteopetrosis [10, 14]. Additionally, cortactin-deficient mice do not have an osteoclast phenotype. These results suggest that unknown proteins are involved in actin ring formation by Src. The cytolinker protein, plectin, was identified as a Src-binding protein. Plectin is a large protein (> 500 kDa) that binds to multiple proteins, including actin and tubulin, and functions as a scaffold for protein-protein interactions [15, 16]. We, first, examined plectin expression by western blotting and found that plectin expression was higher in osteoclasts than in macrophages, that are osteoclast precursor cells [17]. Plectin, along with Src, was localized around the actin ring of osteoclasts [17]. These results suggest that plectin is involved in the organization of the actin ring by Src. Plectin has at least 12 transcriptional variants that differ from each other in exon 1 [16, 18]. The different N-terminal regions of plectin regulate localization in the cytoskeleton [16]. The antibody against plectin that was used in the western blotting and immunofluorescence analyses recognizes pan-plectin and cannot distinguish between the variants [17]. However, qPCR analysis suggested that plectin 1d is higher expressed in osteoclasts than in the skeletal muscle (Figure 1).
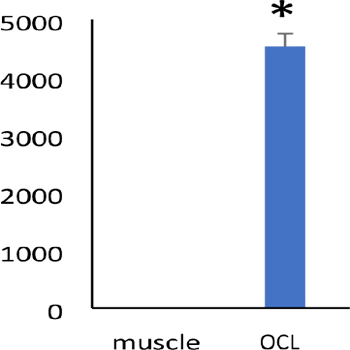
Figure 1: Plectin isoform d was highly expressed in osteoclasts.
mRNAs were isolated from the quadricep muscle (indicated as muscle) and osteoclasts were differentiated from bone marrow cells with M-CSF (20 ng/ml) and RANKL (100 ng/ml) (indicated as OCL). cDNA was synthesized from 1 μg of total RNA by using SuperScript II transcriptase and random primers. Real-time quantitative PCR was performed using cDNA. The data was normalized by actin. * denotes P < 0.01. Representative data from at least two mice are shown for the experiment. Plectin 1d is localized to podosomes and regulates podosome formation [18]. These results suggest that plectin 1d is involved in the actin ring formation of osteoclasts. We, next, examined the interaction between Src and plectin using Halotagged plectin. Plectin bound to Src and was phosphorylated by constitutively activated Src [17]. These results suggest that plectin is one of the proteins downstream of Src in actin ring regulation. The role of plectin was examined by introduction of plectin shRNA in an osteoclast precursor cell line, RAW 264.7. Surprisingly, downregulation of plectin suppressed the differentiation of osteoclasts [17]. During differentiation, osteoclast precursor cells migrate, change shapes (form protrusions), and then attach to each other [5, 19]. Plectin binds to integrins, actin, tubulin, and intermediate filaments, and regulates cell shape and migration [15]. Moreover, plectin functions as a scaffolding platform in various signaling pathways, such as in the MAPK and PKC signaling pathways [15]. RANK-RANKL signaling promotes osteoclastogenesis by activating MAPK signaling and activation of c-fos, p38, and c-Jun/JNK is essential for osteoclastogenesis and for expression and activation of NFATc1 [1, 3, 20]. PKC also plays an important role in osteoclastogenesis through Erk signaling [21]. These results suggest a hypothesis in which plectin functions as a scaffold for MAPK and PKC signaling during osteoclast differentiation. Downregulation of plectin also disturbed actin ring formation in osteoclasts [17]. Src and downstream proteins were activated by integrin signaling, when osteoclasts attached to the bone matrix and organized the actin ring [5]. Plectin has both actin- and integrin-binding domains [16]. These results suggest that plectin plays an important role in actin ring organization through integrin and Src signaling. Taken together, plectin plays an important role in osteoclast differentiation and actin ring formation through Src binding and phosphorylation (Figure 2).
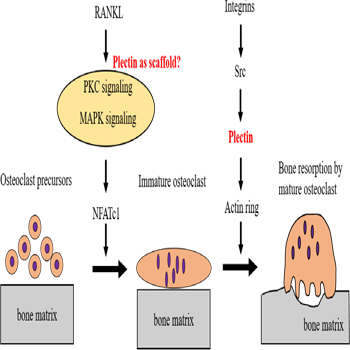
Figure 2: Scheme for the role of plectin in osteoclasts.
Osteoclast precursors are differentiated to osteoclasts by RANKL signaling. Plectin may act as a scaffold for MAPK and PKC signaling and may activate NFATc1, a master regulator of osteoclast differentiation. Plectin also plays an important role after the differentiation of osteoclasts. Plec- tin links integrin and Src signaling to the organization of the actin ring as an actin binding protein.
ACKNOWLEDGEMENTS
Affiliations: This work was supported by grants from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (KAKENHI 25670870 and 16K20423 to T.M., KAKENHI 23249085 to T.T-Y.), and the Fukuoka Foundation for Sound Health Cancer Research Fund to T.M. We would like to thank Editage (www.editage.jp) for English language editing
CONFLICT OF INTEREST
All authors state that they have no conflicts of interest.
REFERENCES
- Takayanagi H. (2007). Osteoimmunology: shared mechanisms and crosstalk between the immune and bone systems, Nat. Rev. Immunol. 7(4): 292-304
- Soysa NS, Alles N, Aoki K and Ohya K. (2012). Osteoclast formation and differentiation: An overview, J. Med. Dent. Sci. 59(3): 65-74.
- IkedaF, Nishimura R, Matsubara T, Tanaka S, et al. (2004). Critical roles of c-Jun signaling in regulation of NFAT family and RANKL-regulated osteoclast differentiation, J. Clin. Invest. 114(4): 475-484.
- Jurdic P, Saltel F, Chabadel A and Destaing O. (2006). Podosome and sealing zone: specificity of the osteoclast model, Eur. J. Cell Biol. 85(3-4): 195-202
- Takahashi N, Ejiri S, Yanagisawa S and Ozawa H. (2007). Regulation of osteoclast polarization, Odontology. 95(1): 1-9.
- Soriano P, Montgomery C, Geske R and Bradley A. (1991). Targeted disruption of the c-src proto-oncogene leads to osteopetrosis in mice, Cell. 64(4): 693-702.
- Boyce BF, Yoneda T, Lowe C, Soriano P, et al. (1992). Requirement of pp60c-src expression for osteoclasts to form ruffled borders and resorb bone in mice, J. Clin. Invest. 90(4): 1622-1627.
- Destaing O, Sanjay A, Itzstein C, Horne WC, et al. (2008). The Tyrosine Kinase Activity of c-Src Regulates Actin Dynamics and Organization of Podosomes in Osteoclasts, Mol. Biol. Cell. 19(1): 394-404
- Duong LT, Lakkakorpi PT, Nakamura I, Machwate M, et al. (1998). PYK2 in osteoclasts is an adhesion kinase, localized in the sealing zone, activated by ligation of alpha (v) beta3 integrin, and phosphorylated by src kinase, J Clin Invest. 102(5): 881-892.
- Nagai Y, Osawa K, Fukushima H, Tamura Y, et al. (2013). p130Cas, Crk-associated substrate, plays important roles in osteoclastic bone resorption, J. Bone Miner. Res.28(12): 2449-2462.
- Nakamura I, Jimi E, Duong LT, Sasaki T, et al. (1998). Tyrosine phosphorylation of p130 (Cas) is involved in actin organization in osteoclasts, J. Biol. Chem. 273: 11144- 11149
- Matsubara T, Myoui A, Ikeda F, Hata K, et al. (2006). Critical role of cortactin in actin ring formation and osteoclastic bone resorption, J Bone Min. Metab. 24(5): 368-372.
- Tehrani S, Faccio R, Chandrasekar I, Ross FP, et al. (2006). Cortactin has an essential and specific role in osteoclast actin assembly, Mol Biol Cell. 17(7): 2882-2895.
- Gil-Henn H, Destaing O, Sims NA, Aoki K, et al. (2007). Defective microtubule-dependent podosome organization in osteoclasts leads to increased bone density in Pyk2 (-/- ) mice, J. Cell Biol. 178(6): 1053-1064.
- Wiche G and Winter L. (2011). Plectin isoforms as organizers of intermediate filament cytoarchitecture, Bioarchitecture. 1(1): 14-20.
- Castañón MJ, Walko G, Winter L and Wiche G. (2013). Plectin-intermediate filament partnership in skin, skeletal muscle, and peripheral nerve, Histochem. Cell Biol. 140(1): 33-53.
- Matsubara T, Kinbara M, Maeda T, Yoshizawa M, et al. (2017). Regulation of osteoclast differentiation and actin ring formation by the cytolinker protein plectin, Biochem. Biophys. Res. Commun. 489(4): 472-476.
- McInroy L and Määttä A. (2011). Plectin regulates invasiveness of SW480 colon carcinoma cells and is targeted to podosome-like adhesions in an isoform-specific manner, Exp. Cell Res. 317(17): 2468-2478.
- Shin N, Choi H, Neff L, Wu Y, et al. (2014). Dynamin and endocytosis are required for the fusion of osteoclasts and myoblasts, J. Cell Biol. 207(1): 73-89
- Matsuo K, Galson DL, Zhao C, Peng L, et al. (2004). Nuclear Factor of Activated T-cells (NFAT) Rescues Osteoclastogenesis in Precursors Lacking c-Fos, J. Biol. Chem. 279: 26475-26480.
- Lee S, Kwak H, Chung W, Cheong H,et al. (2003). Participation of protein kinase c β in osteoclast differentiation and function, Bone. 32: 217-227.